ÔŊâCRISPRģųŌōžŨŧŧĐgĩÄĀûÅcą×
ÕfĩŊģųŌōžŨŖŦ´ķŧŌļŧūĪëÆđCRISPRŧŧĐgĄŖß@ŽÄęÃû´ķÔëĩÄŧŧĐgŖŦŦFŊņŌĀÅfáļȲģpŖŦĶČÆäĘĮÎŌĩÄCRISPR´ķÉņähŖŦŊüÆÚ°ląíĩÄÎÄÕÂîlîlÁÁĪāĶÚÖĒÃûësÖžŖŦĶÖŌũÆđŌģÆŦá×hĄŖrß^žŗßwŖŦCRISPRŧŧĐg˛ĸ]ĶĐäNÂÄäÛEŖŦŌģÖąÔÚžĄŖĩĢČÎēÎĘÂÎī°üĀ¨ŧŧĐgĶĐĀûžÍĶĐą×ŖŦCRISPRŧŧĐgŽČģŌ˛ēÁ˛ģĀũÍâĄŖ
CRISPRŧŧĐgžÍĘĮß@ôēÃĶÃŖĄ
ģđÁËËÄÄęÖŽžÃĩÄŧŧĐgŖŦļúĘėÄÜÔĩÄŨī@ļøŌ×ŌŖēŪđÁËĻĶÚESŧ°ûĩÄŌĀŲŖŦCRISPRŧŧĐgŦFÁËĻ´ķĘķĄĸØiĄĸēīĄĸ°ßņRô~ĩČļāˇNÎīˇNĩÄžŨŖģCRISPRŧŧĐgŌĀŲĶÚCas9ēÍsgRNAŦFĄ°žĢĘĄąĮиîŖŦēģ¯ō˛ŊķEŖŦ´ķ´ķŋsļĖōÖÜÆÚĄŖÖÜÆÚļĖĄĸš¤×÷ÁŋĐĄŖŦëSÖŽļøíĩÄžÍĘĮŲMĶÃĩÄŊĩĩÍĄŖžCÉĪŖŦĐÔrąČ¸ßĩÄŧŧĐgŽČģŽČʲģ×ĩÄūŌģÖąģđĄŖ

CRISPRŧŧĐgŌ˛ĶĐČąücŖŋ

CRISPRŧŧĐg×ÔĘĀŌÔíŖŦąģĒĶÃÖÁĘĀŊį¸÷ĩØąļāōĘŌÖĐßMĐĐŋÆWŅĐžŋŖŦÔÚĪíĘÜÆäŨĩÄÍŦrŖŦËüĩÄą×ļËŌ˛ĘĮąËųÖÜÖĒŖēsgRNAÅcCas9ĩ°°×ß@¸įzŖŦ˛ĸ˛ģĘĮrrŋĖŋĖļŧž¤ž¤IIš¤×÷ĩÄŖŦÕlß]ĶĐžëĩĄĩÄrēōÄØŖŋąžĒÔžĢĘļ¨ÎģĩÄsgRNAŌ˛ĶĐ×ßŅÛĩÄrēōŖŦąžĒÔ×ReËĘPAMĐōÁĐ ĩÄCas9ĩ°°×ŖŦŌ˛Åŧ ˛ģ×ßĐÄŖŦŊYšûžÍĘĮðĐŖŦĶ°íÁËžŨĩÄЧÂĘĄŖ
ĶĐĄ°˛ĄĄą˛ģŋÉÅÂŖŦęPæIĘĮĄ°Ļ°YĪÂËĄąŖŋ
átÉúÖβĄÖvžŋĻ°YĪÂËŖŦŋÆŅĐËŧ¡Ō˛˛ģĀũÍâĄŖÕŌŗöÔŌōŊâQî}ŖŦŧČČģĘĮsgRNAēÍCas9ĩ°°×ŗöŦFÁËÃĢ˛ĄŖŦÄĮôÎŌžÍáĻ˛ģÍŦ˛Ą°Y˛ÉČĄ˛ģÍŦˇŊĘŊŖē

ģ¯sgRNA
Ŗ¨1ŖŠ¸Ä××ÔÎŌŖē¸Ä×sgRNA ĩÄéLļČŖŦŊØā5' ļËĩÄ2-3 AģųģōÔÚ×ReĐōÁĐĮ°ŧĶ2 øBāŅßĘŖŦĮиîģîĐÔ˛ģ×ŖŦ ðĐÂĘŊĩĩÍ5000 ąļĄŖ
Ŗ¨2ŖŠĖáÉũ×ÔÎŌŖēÔöŧĶsgRNA ĩġļ¨ĐÔĄŖČįÔÚgRNA ÖĐÔöŧĶŌģĻA-U AģųÅäĻĩȡŊˇ¨ĄŖ´ËÍâŖŦ ĶĐŅĐžŋąíÃ÷Cas9 ÖąĪĩÍŦԴøĻÄŗĐŠÎģücĩÄĖØŽĐÔŨ^SpCas9 ¸ßŖŦß@oŌÉéĖá¸ßĖØŽĐÔĖᚊÁËŌģlĐÂËŧ¡ĄŖ
Ėá¸ßCas9ĩ°°××ÔÉíĖØŽĐÔ
ŖŌģĐÔĻĶÚŧŌÍĨĘĮēÜÖØŌĒŖŦļøĻĶÚCRISPRŧŧĐgŌ˛ĘĮēÜÖØŌĒĄŖŖŌģĐÔŌģˇŊÃæÅc×ÔÉíĶĐęPŖŦÁíŌģˇŊÃæŌ˛ÅchžŗĩÄÕTģķĶĐęPŖŦŌō´ËĻĶÚCas9ĩ°°×ŖŦÎŌŌĒÄČēÍÍâÉŌōËØČĨŋŧ]ĄŖ

Ŗ¨1ŖŠ ˛ÁÁÁŅÛžĻŖŦąæÕJŗöÕlĘĮÄãĩÄMr/Mrs RightŖēÔöCas9 ĩ°°×ĻËĘPAM ĐōÁĐĩÄ×ReÄÜÁĻŖŦÍģ×ķwD1135EĻËĘPAM ĩÄ×Re¸üŧĶĖØŽŖŦÄļøŊĩĩÍŌō×ReˇĮËĘPAM ĐōÁĐļøŌũÆđÃ°ĐĄŖ
Ŗ¨2ŖŠÅcĻĩÄČËĪāĶHĪāÛžÍēÃŖŦ˛ģŌĒĀĪĘĮÕ´ģ¨ČĮ˛ŨŖēĸCas9ĩ°°×ßMĐĐļ¨ücÍģ×ĐÎŗÉeSpCas9ēÍSpCas9-HF1ŖŦÍģ×ķwpČõÁËCas9 ĩ°°×ÅcDNA ĩġĮĖØŽĐÔŊYēĪĩÄÄÜÁĻŖŦÔö°ĐĪōĐōÁĐÅcCas9 ĩ°°×ŊYēĪĩĸ ÁĻĄŖ

Ŗ¨3ŖŠpÉŲÍâŊįÕTģķŖēÕ{ŋØCas9 ĩ°°×ÔÚŧ°ûÖĐĩÄąíß_ÁŋĄŖͨß^ĘšĶÃÕT§ĐÍĸĶ×ĶĄĸ˛åČë4-HT ŌĀŲĩÄČēŦëÄĩȡŊˇ¨Ŋ¨ÕT§ĐÍCas9 ĩ°°×ŖŦpÉŲģųŌōŊMąŠÂļĶÚCas9 ÅcsgRNA ÍēĪķwĩÄrégŖŦ ŧ´pÉŲCas9 ÅcˇĮ°ĐĪōĐōÁĐĩÄŊYēĪ¸ÅÂĘŖŦßMļøŊĩĩÍðĐÂĘĄŖ
Ŗ¨4ŖŠāÉáëxŖēHąŖÁôÆäēËËá×ReÄÜÁĻŖŦāŗũÆäēËËáČĮĐøĩÄģîĐÔĄŖŽČģĻĶÚ×ÔÉíŌ˛ĘĮąØíĩÄŖēŊĩĩÍHNH ģōRuvC ŊYĶōšĻÄÜĩÄÍģ×ķwH840A ēÍD10AŖģŊ¨ĘšCas9 ĩ°°×ĘʧēËËáČĮĐøģîĐÔŖŦŊčÖúÆäËûēËËáČĮĐøŦFģųŌōžŨĩÄCRISPRiēÍFokĸņ-dCas9ĄŖ
°ŲWŲDĸĘÖīĩĄĒĄĒSouthern blotzyŊđËĘ

sgRNAÔOĶĀÖ
ÉáŋÉÄÜðĐĩŊĶĐšĻÄÜģųŌōŊMĐōÁĐĩÄsgRNA
ŧRNA×ĸÉä
ŋsļĖCas9ĩ°°×ēÍsgRNAĩÄ×÷ĶÃrég
Southern blotzy
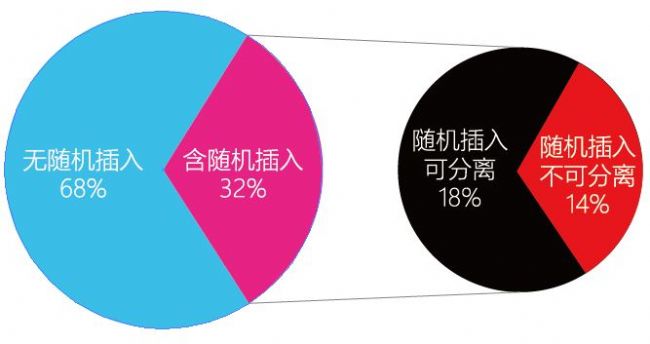
Ļ´ķÁŋĩūŊyĶĩūˇÖÎö°lŦFŖŦ˛ÉĶÃESŧ°ûˇŊĘŊÖÆäŖŦëSC˛åČë¸ÅÂĘŧsé20%ŖŦ˛ÉĶÃCRISPR/Cas9ŧŧĐgŖŦ´ķŧsĶĐ32%ĩĸÅÂĘĶĐëSC˛åČëĄŖļøß@32%ÖĐĶĐ14%ĩÄëSC˛åČëoˇ¨ŌĀŋŋŊģÅä÷´úČĨŗũŖŦÅÅŗũëSC˛åČëĩÄŊđËĘĘĮSouthern blotĄŖ°ŲWŲDĀ°ŅŲ|ÁŋęPŖŦÔŗÖßMĐĐSouthern blotzyŖŦ´_ąŖģųŌōžŨĢ@ĩÃĩÄĶÎīoëSC˛åČëĄŖ
SouthernësŊģĩÄÄŋĩÄĘĮéÁËzyÄŋĩÄģųŌōĩIJåČëÎģÖÃŧ°ŋŊØĩÁŋĄŖSouthern blotësŊģĘĮßMĐĐģųŌōŊMDNAĖØļ¨ĐōÁĐļ¨ÎģĩġŊˇ¨ŖŦÆä˛Ų×÷ß^ŗĖŖēĀûĶÃëĶžˇÖëxŊøĪûģ¯ēķĩÄDNAÆŦļÎŖŦČģēķÔŲĸÄzÉĪĩÄDNA×ĐÔ˛ĸÔÚÔÎģĸÎæDNAÆŦļÎŪDŌÆÖÁÄáũĤģōÆäËûšĖĪāÖ§ŗÖÎīÉĪŖŦŊ¸ÉŋžģōÕß×ĪÍâžÕÕÉäšĖļ¨ŖŦÔŲÅcĪāĻĒŊYĩÄDIGËĶĖŊáßMĐĐësŊģŖŦĶÃˇÅÉä×Ôī@Ķ°ģōøˇ´Ēī@ÉĢŖŦÄļøzyĖØļ¨DNAˇÖ×ĶĩÄēŦÁŋĄŖ
ĸŋŧŖē
[1] Cho S W, Kim S, Kim Y, et al. Analysis of off-target effects of CRISPR/Cas-derived RNA-guided endonucleases and nickases[J]. Genome research, 2014, 24(1): 132-141.
[2] Kleinstiver B P, Prew M S, Tsai S Q, et al. Engineered CRISPR-Cas9 nucleases with altered PAM specificities[J]. Nature, 2015, 523(7561): 481.
[3] Terao M, Tamano M, Hara S, et al. Utilization of the CRISPR/Cas9 system for the efficient production of mutant mice using crRNA/tracrRNA with Cas9 nickase and FokI-dCas9[J]. Experimental animals, 2016, 65(3): 275-283.
[4] M¨šller M, Lee C M, Gasiunas G, et al. Streptococcus thermophilus CRISPR-Cas9 Systems Enable Specific Editing of the Human Genome[J]. Molecular Therapy, 2016, 24(3):636¨C644.
[5] Qi L, Larson M, Gilbert L, et al. Repurposing CRISPR as an RNA-Guided Platform for Sequence-Specific Control of Gene Expression[J]. Cell, 2013, 152(5):1173.
[6] Luke A. Gilbert, Matthew H. Larson, Morsut L, et al. CRISPR-Mediated Modular RNA-Guided Regulation of Transcription in Eukaryotes[J]. Cell, 2013, 154(2):442.
[7] Hsu P D, Scott D A, Weinstein J A, et al. DNA targeting specificity of RNA-guided Cas9 nucleases[J]. Nature Biotechnology, 2013, 31(9):827-32.
[8] Fu Y, Sander J D, Reyon D, et al. Improving CRISPR-Cas nuclease specificity using truncated guide RNAs[J]. Nature Biotechnology, 2014, 32(3):279.
[9] Kleinstiver B P, Vikram P, Prew M S, et al. High-fidelity CRISPR-Cas9 variants with undetectable genome-wide off-targets:[J]. Nature, 2016, 529(7587):490-495.
[10] Slaymaker I M, Gao L, Zetsche B, et al. Rationally engineered Cas9 nucleases with improved specificity[J]. Science, 2016, 351(6268):84.
[11] Chiang T W W, Sage C L, Larrieu D, et al. CRISPR-Cas9 D10A nickase-based genotypic and phenotypic
screening to enhance genome editing[J]. Scientific Reports, 2016, 6:24356.
[12] Pan Y, Shen N, Jungklawitter S, et al. CRISPR RNA-guided FokI nucleases repair a PAH variant in a phenylketonuria model[J]. Scientific Reports, 2016, 6:35794.
ÖÆ×÷Ŗē
MITû¸ęÎÄÄXŋÆWŅĐžŋËų
äh
SputnikĶŽ

- ÖØcŨášâČĢøÕĐÄŧÖ˛Îīßz÷ŪDģ¯ÅcģųŌōžŨˇūÕŊäNÉĖ
- ŨášâÉúÎīÍÆŗöģųŌōžŨŨdķwŊ¨ÅcÖ˛Îīßz÷ŪDģ¯ˇūÕ
- °ŲȤÉúÎīŖŦQPCRzyĩÍríŌuŖŦÚAĄ°0ÔĒŌũÎīēĪŗÉĄą
- Esco Ventures X Ą°ģųŌōÖίēąŌ˛ĄĄąžWÉĪÕ¯
- ÕnŗĖîA¸æŖŦÄĪÄŖÉúÎīōĘŌĐĄĘķĒĶÃŧ°˛Ų×÷ĪĩÁĐÖv×ų
- ÄĪˇŊÄŖĘŊÉúÎīÕ\ƸļāĩØ ^ŋÆŅĐäNĘÛžĢĶĸ
- ÄĪÄŖÉúÎīCre-loxPĪĩŊyŊéŊBŧ°ĘšĶÃî}Öv×ųŅûÕ
- ŲIÉúÎīŗÉšĻÍęŗÉ2.85|ČËÃņÅBŨČÚŲY