
m6A修飾在家畜生長發(fā)育和繁殖性狀中的調控作用研究進展
通過甲基轉移酶對真核RNA的N6-甲基腺苷(m6A)進行可逆調控是影響RNA代謝的一種重要表觀遺傳事件。因此,m6A甲基化在調控動物生長、發(fā)育、繁殖和疾病進展中發(fā)揮著至關重要的作用。本文回顧了m6A甲基化修飾的最新研究進展,并討論了其在家畜生長、發(fā)育和繁殖性狀中的調控作用,展望了m6A甲基化修飾在塑造經濟重要性狀中的研究前景。
m6A的基本調控機制
m6A修飾是一種常見的RNA修飾類型,其可以改變RNA分子的化學屬性和空間結構,從而影響其功能性。m6A修飾動態(tài)可逆,參與多種酶之間的復雜互作,包括甲基轉移酶(如METTL3、METTL14和WTAP)、去甲基化酶(如FTO、ALKBH5和ALKBH3)以及m6A結合蛋白(如YTHDF1、YTHDF2、YTHDF3、YTHDC1和YTHDC2)。研究表明,m6A修飾可以影響mRNA的剪接、穩(wěn)定性、出核和翻譯,以及miRNA的加工和功能(圖1) 。
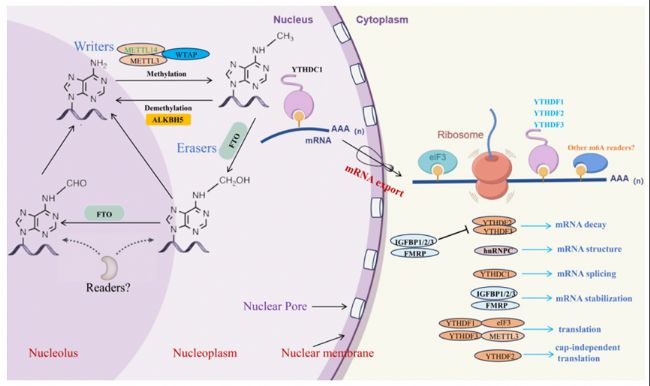
圖1:m6A甲基化修飾的動態(tài)過程。負責添加m6A修飾酶是蛋白質復合體,包括 METTL3、METTL14、WTAP 和其他組分。FTO蛋白是一種去甲基化酶,通過其核心結構域C末端的獨特long loop區(qū)域作用,以去除RNA中的甲基化。ALKBH5是另一種去甲基化酶,通過氧化過程去除帶有m6A修飾的RNA的甲基化。諸如YTHDF1、YTHDF2、YTHDF3、YTHDC1和YTHDC2等reader蛋白參與了包括mRNA剪接、穩(wěn)定性、翻譯和出核在內的各種過程。
m6A 修飾在動物生長發(fā)育中的作用
(1)m6A修飾在肌肉發(fā)育中的調控作用
作為最豐富的mRNA修飾類型,m6A在骨骼肌分化、心肌重塑和再生中發(fā)揮著調控作用。在豬胚胎發(fā)育的6個不同階段中,m6A修飾顯示出高度動態(tài)變化,大多數受影響的基因富集在與骨骼肌發(fā)育相關的通路中。豬產前骨骼肌中的m6A甲基化水平在胚胎年齡E40(原始肌纖維形成期)時顯著降低,隨著胚胎年齡增加(E50-95天,次級肌纖維形成期),甲基化水平逐漸增加。通過siRNA沉默IGF2BP1可以促進增殖并抑制肌細胞分化。骨骼肌發(fā)育在家畜高效肉類生產中的核心地位突顯了m6A修飾對產品質量的重要性。山羊研究表明,FTO可以通過降低m6A修飾程度來增加GADD45B mRNA的穩(wěn)定性和表達,從而激活p38MAPK通路并促進肌細胞分化。METTL3蛋白也可以通過上調m6A水平和增強MEF2C表達來促進山羊骨骼肌衛(wèi)星細胞的肌源性分化。這些研究為通過RNA甲基化調控山羊肌生成提供了潛在的分子機制。
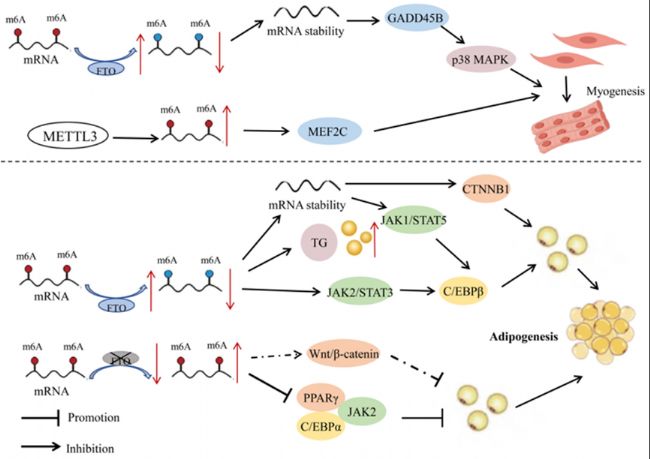
圖2:RNA甲基化在家畜肌肉和脂肪發(fā)育中的機制。在肌細胞中,FTO通過降低m6A修飾程度以增加GADD45B mRNA的穩(wěn)定性和表達,從而激活p38MAPK通路,促進肌細胞分化。另一方面,METTL3通過上調m6A水平和增強MEF2C表達,促進山羊骨骼肌衛(wèi)星細胞的分化。在脂肪細胞中,FTO通過去甲基化調控JAK2-STAT3-C/EBPβ信號通路,促進豬前脂肪細胞分化。FTO過表達還可以顯著增加細胞中的甘油三酯(TG)含量。此外,FTO通過調控CTNNB1表達促進雞前脂肪細胞增殖和分化。沉默FTO顯著降低PPARγ和C/EBPα表達,抑制豬肌內前脂肪細胞的增殖。它還可能通過Wnt/β-catenin通路參與豬肌內前脂肪細胞分化。在豬骨髓源性間充質干細胞(BMSCs)中,METTL3缺失增加了JAK1 mRNA穩(wěn)定性,通過JAK1-STAT5信號通路激活C/EBPβ,從而誘導BMSCs分化為脂肪細胞。紅色和藍色圓點分別代表高甲基化和低甲基化狀態(tài)。虛線表示潛在的調控機制。符號:↑表示上調;↓表示下調。
在對牛的研究中,m6A甲基化轉移酶METTL3對成肌細胞增殖具有抑制作用,并能促進細胞分化;它還能通過影響TM4SF1穩(wěn)定性來調控成肌細胞發(fā)育。YTHDF2介導的TM4SF1 mRNA降解也依賴于m6A修飾。成年牛背最長肌中m6A去甲基化酶(FTO和ALKBH5)的表達顯著高于新生牛,而m6A甲基轉移酶(METTL3、METTL14和WTAP)的表達則較低。在肌生成過程中,這五個基因的mRNA表達顯著增加;在牛成肌細胞中的干擾實驗進一步表明,m6A甲基化相關酶可能調控牛骨骼肌發(fā)育。
在京星黃(Jingxing-Huang)雞中,m6A去甲基化酶基因ALKBH5和FTO在骨骼肌發(fā)育過程中的表達水平顯著增加。ALKBH5和FTO蛋白可能通過調控RNA m6A甲基化水平影響雞骨骼肌發(fā)育。有證據表明METTL3、METTL14和WTAP在家禽肌纖維類型組成、維持和成肌細胞分化中具有調控作用。
m6A是影響非編碼RNA(ncRNA)功能的關鍵調控因子,從而參與生物體的生長和發(fā)育。METTL3過表達降低了分化C2C12細胞和肌肉損傷再生小鼠模型中肌肉特異性miR-1a、miR-133a、miR-133b和miR-206表達。研究人員使用RNA-seq和MeRIP-seq技術分析了豬氧化和糖酵解骨骼肌中l(wèi)ncRNAs的m6A甲基化模式。結果顯示,大多數lncRNAs在最后一個外顯子中有一個優(yōu)先富集的m6A peaks,且m6A水平與lncRNAs表達水平之間存在正相關。此外,lncRNA MSTRG.14200.1的抑制延遲了衛(wèi)星細胞分化,并刺激了快速和慢速肌纖維之間的轉換。這項研究為豬肌纖維類型的轉換提供了新的目標。在牛-牦牛骨骼肌中,m6A豐度與基因表達水平呈正相關,但與lncRNA表達水平呈負相關,表明m6A修飾可能在牛-牦牛骨骼肌的發(fā)育中發(fā)揮重要作用,但m6A修飾對mRNA和lncRNA的調控機制可能不同。RNA-seq和ATAC-seq測序技術的使用允許發(fā)現骨骼肌特異性lncRNA MYH1G-AS,其轉錄由轉錄因子SMAD3和SP2協(xié)調。此外,SP2抑制ALKBH5的轉錄,降低了ALKBH5對MYH1G-AS的去甲基化,從而MYH1G-AS RNA穩(wěn)定性失衡。MYH1G-AS加速了成肌細胞的增殖,但抑制其分化;這個基因促進了慢速肌纖維向快速肌纖維的轉換,并導致肌肉萎縮。總體而言,這些一致的結果表明m6A甲基轉移酶在家畜骨骼肌發(fā)育中發(fā)揮重要的調控作用。
(2)m6A修飾在脂肪生成中的調控
有證據表明m6A修飾可以調控轉錄因子和脂肪組織特異性基因的表達水平,從而影響脂肪沉積代謝過程。其中,FTO作為一種RNA去甲基化酶,在修飾基因表達中發(fā)揮重要作用。例如,在豬脂肪細胞中FTO和METTL3基因的過表達和特異性敲除揭示了FTO與m6A甲基化水平呈負相關,而與脂肪生成呈正相關。而METTL3與m6A甲基化水平呈正相關,與脂肪生成呈負相關。在豬骨髓源性間充質干細胞(BMSCs)中,METTL3缺失導致JAK1 mRNA上的m6A修飾減少,進而增強了JAK1 mRNA的穩(wěn)定性和翻譯效率。JAK1-STAT5信號通路是調節(jié)脂肪細胞分化的關鍵通路之一,通過磷酸化STAT5激活與脂肪生成相關的下游基因,如C/EBPβ,并促進其進入細胞核,從而引導BMSCs向脂肪細胞分化。FTO酶通過m6A去甲基化促進豬的前脂肪細胞分化,通過調節(jié)JAK2-STAT3-C/EBPβ信號通路、有絲分裂克隆擴增和細胞自噬通路。在豬肌內前脂肪細胞中,沉默FTO顯著降低了磷酸組蛋白H3水平,并抑制肌內前脂肪細胞增殖。沉默FTO還導致過氧化物酶體增殖物激活受體γ(PPARγ)和CCAAT/增強子結合蛋白α(C/EBPα)下調,以及β-catenin表達的上調。LiCl(一種Wnt/β-catenin信號通路的特異性激活劑)可以減弱FTO誘導的PPARγ上調和β-catenin下調。在豬脂肪細胞中,FTO過表達顯著增加了細胞內甘油三酯含量,并顯著上調了與脂肪生成相關基因的C/EBPβ表達,同時下調了脂肪分解基因ATGL和HSL的mRNA表達;FTO敲低會導致不同結果,表明這種蛋白可能促進豬脂肪細胞中脂質的積累。總之,沉默 FTO 會減少豬肌內脂肪前細胞的增殖和分化,并可能通過 Wnt/β-catenin 通路影響豬肌內脂肪前細胞的分化。
在中國本土雞種中,腹部脂肪過度沉積一直是家禽生產中的一個重大問題。高脂飲食可能導致體重增加和過度脂肪沉積,影響脂肪代謝和脂肪生成的基因和通路。在雞的前脂肪細胞中,RNA m6A甲基化與脂肪生成呈負相關。FTO過表達顯著抑制m6A水平,并促進雞的前脂肪細胞的增殖和分化,而沉默FTO則產生相反結果。FTO酶通過去甲基化調控CTNNB1表達,從而促進雞的脂肪沉積。有研究報告稱,與新生牛相比,秦川成年牛脂肪組織中的m6A甲基轉移酶METTL3、METTL14和WTAP以及m6A去甲基化酶FTO和ALKBH5都顯著上調。它們在體外前脂肪細胞增殖和分化過程中的表達模式也表現出顯著差異。這些結果表明m6A修飾可能在家牛的脂肪沉積中發(fā)揮重要作用,m6A甲基轉移酶在調控牛的前脂肪細胞的增殖和分化中具有潛在作用。
當前研究表明,許多環(huán)狀RNA(circRNA)分子具有多個m6A修飾位點。例如,在黔山黑豬和大白豬的背最長肌中鑒定出12個候選m6A修飾circRNA(m6A-circRNA);這些m6A-circRNA可以作為miRNA海綿分子,并通過構建ceRNA調控網絡來調控脂肪沉積。富集分析顯示,m6A-circRNA的親本基因及其吸附的miRNA靶基因參與與脂肪沉積、細胞增殖和分化相關通路,表明m6A-circRNA在決定豬肉品質方面可能特別重要。這些一致的發(fā)現表明,m6A甲基轉移酶在家牛、豬和家禽的脂肪沉積中發(fā)揮著至關重要的作用。
m6A修飾在家畜繁殖性狀中的功能性作用
(1)m6A修飾對精子發(fā)生的調控
精子發(fā)生是一個高度復雜和特殊的細胞發(fā)育過程,包括精原細胞增殖、精母細胞減數分裂和精子形成三個階段。精子發(fā)生過程中的基因表達在轉錄、轉錄后和翻譯水平上受到嚴格調控,以確保階段特異性基因的準確表達。以往的研究使用m6A RNA甲基轉移酶METTL3和/或METTL14的條件性基因敲除小鼠模型揭示了METTL3或METTL14在早期精原細胞中的缺失會導致精原干細胞凋亡。然而,只有在相對較晚階段的精原細胞(A1型精原細胞)中同時敲除METTL3和METTL14,才會影響精子形成。在人類中,WTAP和ALKBH5表達在精子發(fā)生過程中減少;FTO和ALKBH5在哺乳動物的睪丸中高表達,人類FTO基因突變與精液質量下降密切相關。有研究報告稱,ALKBH5基因缺陷的雄性小鼠由于減數分裂中期精原細胞凋亡而生育能力降低。山羊睪丸發(fā)育中ALKBH5表達上調,山羊精原干細胞中ALKBH5過表達促進了從G1期到S期轉變并抑制凋亡。研究人員使用m6A-seq、RNA-Seq和核糖體分析技術,生成了精子發(fā)生不同發(fā)育階段m6A RNA修飾的動態(tài)圖譜。研究結果表明m6A甲基轉移酶失活導致m6A RNA修飾水平下降,從而導致參與精原細胞命運決定和精子形成的基因轉錄和翻譯效率發(fā)生顯著變化。
轉錄延伸標記H3K36me3介導m6A修飾的共轉錄過程。H3K36me3與m6A甲基轉移酶復合體中的METTL3和METTL14互作,促進新生RNA m6A修飾。組蛋白和RNA修飾之間的這種交流為mRNA-m6A的生物發(fā)生提供了新的概念。盡管DNA甲基化和m6A修飾是獨立過程,但它們可能共同影響基因表達和精子發(fā)生的調控。非編碼RNA作為表觀遺傳調控因子,在精子發(fā)生過程中起著重要作用。其中,miRNA、circRNA和PiRNA在轉錄、轉錄后和翻譯水平上調節(jié)雄性生殖細胞的正常發(fā)育,并可能與m6A修飾互作。先前的研究表明,在小鼠GC-1精原細胞中,敲低METTL3后,構建的circRNA-miRNA-mRNA調控網絡揭示了H2afx和Dnmt3a是精子減數分裂中的樞紐基因。這項研究可能為由異常m6A修飾引起的異常精子發(fā)生的病理機制提供新的見解和潛在的治療靶點。
豬生殖細胞中的所有mRNA轉錄本都含有m6A修飾,m6A的reads密度在編碼序列(CDS)中增加,并在終止密碼子附近達到最高值,然后在3'非翻譯區(qū)(UTR)中降低。在精子發(fā)生過程中,轉錄產物中m6A修飾的豐度隨著不同發(fā)育階段而變化。粗線期精母細胞和圓形精子細胞中CDS的m6A修飾reads密度較高,而在精原細胞的30非編碼區(qū)中較高。METTL3缺失會導致精原細胞分化異常啟動,并影響精母細胞在減數分裂前達到粗線期的能力。在粗線期精母細胞中,甲基化基因上調優(yōu)先參與精子發(fā)生和細胞周期過程,這些過程被m6A修飾上調,表明m6A在早期發(fā)育階段的重要作用。此外,PS中的m6A修在參與精子細胞發(fā)育基因中富集,并上調參與微管加工、纖毛組織和纖毛組裝的基因表達,表明m6A甲基化mRNA可能在精子發(fā)生前抑制翻譯。在圓形精子細胞中,m6A甲基化基因上調,并調控包括多細胞生物解剖結構和管狀形態(tài)發(fā)生在內的發(fā)育過程,表明m6A修飾在介導豬精子發(fā)生中的保守性。在雄性生殖細胞發(fā)育過程中的m6A RNA修飾保守并受動態(tài)調控,廣泛存在于精子發(fā)生的各個階段,特別是在粗線期/雙線期精母細胞和圓形精子細胞中高度富集。在牛、牦牛和牦牛-牛雜交品種的睪丸組織中,bta-miR-200a通過抑制ALKBH5的表達調控m6A甲基化水平,從而影響m6A靶基因的表達和功能。這些一致的結果表明m6A甲基轉移酶可能參與動物精子發(fā)生(圖3)。
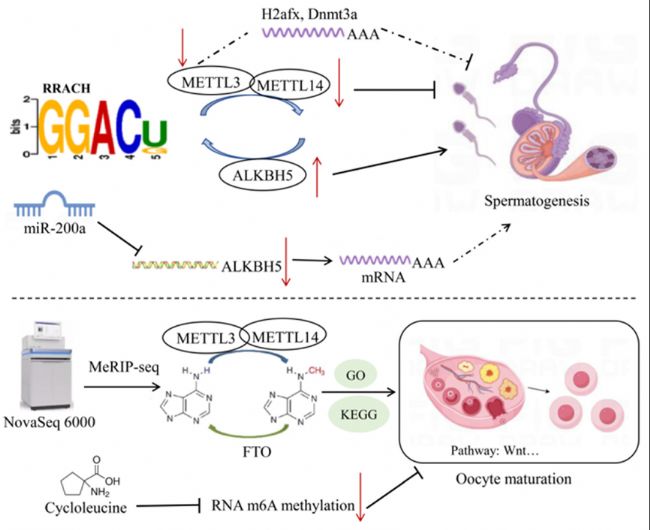
圖3:家畜繁殖性狀中RNA甲基化機制。小鼠GC-1精原細胞中敲低METTL3后,蛋白-蛋白互作(PPI)網絡分析顯示核心基因H2afx和Dnmt3a可能在精子發(fā)生中發(fā)揮作用。在牛睪丸組織中,bta-miR-200a抑制ALKBH5表達,從而影響m6A甲基化對精子發(fā)生相關基因的調控。在豬卵母細胞中,環(huán)亮氨酸抑制m6A甲基化,從而抑制減數分裂和早期胚胎發(fā)育。
(2)m6A修飾對動物卵母細胞成熟的調控
動物卵母細胞成熟可以分為三個主要階段:減數分裂成熟、細胞質成熟和表觀遺傳成熟。卵母細胞的表觀遺傳成熟主要包括調控基因表達的DNA甲基化和組蛋白修飾。m6A甲基化修飾可以影響mRNA降解速率和轉錄本長度,并阻礙正常的mRNA翻譯。這可能導致卵泡發(fā)育缺陷、卵母細胞成熟抑制、卵母細胞早期發(fā)育停滯和合子基因組激活受損,從而影響和調控雌性哺乳動物的卵母細胞形成各個階段。
卵母細胞支持胚胎發(fā)育能力在卵泡生成過程中獲得,與卵泡發(fā)育過程中周圍顆粒細胞的互作可以影響卵母細胞質量。豬顆粒細胞表現出豐富的m6A基因修飾,從小卵泡(<3毫米)到大卵泡(>5毫米)的轉變過程中,m6A甲基化發(fā)生動態(tài)變化。在小卵泡和大卵泡的顆粒細胞中分別鑒定出7289和6882個m6A位點,m6A在5'UTR或3'UTR附近富集并共有保守motif。進一步分析顯示,差異表達的m6A甲基化基因在與類固醇生成、顆粒細胞增殖和卵泡發(fā)育相關的多個信號通路中顯著富集。總體而言,這些結果突出了豬卵泡發(fā)育過程中顆粒細胞中存在不同的m6A修飾,這可能與類固醇和卵泡生成有關。
牦牛的發(fā)情周期是一種季節(jié)性現象,可能涉及性激素合成和分泌以及對卵泡生長和發(fā)育的表觀遺傳調控。HE染色顯示,在發(fā)情期卵巢中的發(fā)育中卵泡和成熟卵泡數量明顯多于黃體期和妊娠期。qPCR結果顯示,在卵巢發(fā)育的不同階段,METTL3、METTL14、FTO和YTHDC1的表達存在顯著差異,表明m6A修飾在卵巢活動中具有調控作用。通過MERIP-seq技術繪制的牦牛m6A轉錄組譜顯示,在編碼序列(CDS)、3'UTR和保守序列“DRACH”中m6A peaks高度富集。GO、KEGG和GSEA分析結果表明,m6A修飾可能參與牦牛卵巢在生殖周期中的許多生理活動。
在家禽研究中,卵泡內的體細胞決定了卵泡的選擇過程。在雞的卵泡選擇過程中對m6A甲基化水平分析顯示,顆粒細胞和卵泡膜細胞中mRNA甲基化和去甲基化廣泛存在,m6A甲基化修飾與基因表達水平呈負相關。在卵泡選擇過程中,m6A甲基化peaks和修飾轉錄本均增加,導致與卵泡生成相關的許多基因表達動態(tài)變化。功能富集分析表明,Wnt通路中關鍵因子的m6A修飾可能在雞的卵泡選擇中發(fā)揮主要作用。
先前研究表明,用m6A抑制劑(環(huán)亮氨酸)處理卵母細胞可以抑制RNA m6A甲基化和組蛋白修飾,并改變囊胚階段代謝基因表達,從而抑制豬卵母細胞的減數分裂和早期胚胎發(fā)育。這些一致的結果表明,m6A甲基轉移酶可能參與主要家畜物種卵母細胞的成熟過程。
總結和觀點
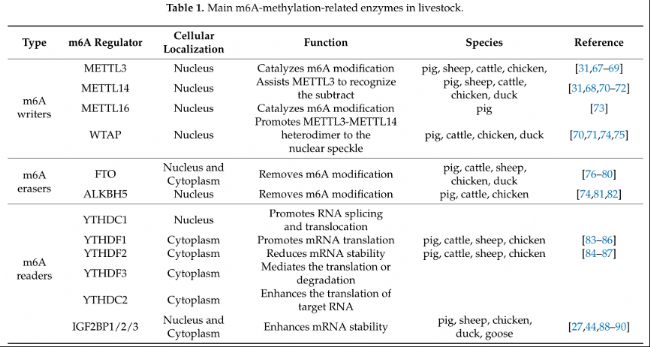
m6A修飾,作為一種廣泛存在的重要表觀遺傳修飾,在農業(yè)動物的各種生命過程中扮演著重要角色,包括生長和發(fā)育、脂質代謝和繁殖(表1)。
近年來,多組學整合分析已成為生命科學領域最重要的研究方法之一。越來越多的研究關注m6A修飾與其他組學數據的綜合分析。例如,轉錄組與m6A之間的相關性可以全面解釋轉錄因子對基因表達和表型差異的調控作用。在子宮內膜癌研究中,通過MeRIP-seq、RNA-seq測序和實驗分析,發(fā)現IGF2BP1鑒定目標基因PEG10終止密碼子附近的m6A甲基化位點,從而調控子宮內膜癌細胞增殖。此外利用全基因組、RNA-seq和功能研究,揭示IGF2BP1基因啟動子區(qū)域缺失可能是與雞體型相關數量性狀位點的因果變異。
與人類和小鼠的m6A修飾研究相比,家畜的m6A修飾研究相對較少。目前,使用m6A-seq評估m(xù)6A修飾在家畜經濟性狀中的作用的研究主要集中在甲基轉移酶、m6A水平以及與性狀相關的差異甲基化基因鑒定上。因此,需要進一步研究家畜中RNA甲基化相關酶和甲基化基因的具體功能和調控機制。RNA甲基化研究仍面臨許多挑戰(zhàn),如m6A-seq測序技術限制可能導致的假陽性結果,以及缺乏對家畜RNA甲基化動態(tài)性質和調控機制的研究。
m6A修飾通常被視為基因表達的調控因子和許多調控通路中的目標。m6A的writers、erasers、readers經常相互交互。在癌癥研究中,METTL3可能調控WTAP蛋白穩(wěn)定性,而WTAP蛋白上調只有在METTL3存在時才具有致癌作用。參與不同類型癌癥的m6A相關蛋白通常受到多種m6A蛋白因子調控。由于對m6A了解有限,仍需要廣泛研究通路之間的互作以及m6A在調控基因表達中的協(xié)同效應。迄今為止,已經發(fā)現了多種writers,但只鑒定出兩種erasers:FTO和ALKBH5。FTO或ALKBH5的異常表達只會導致生物體整體m6A修飾水平的微小變化,這表明體內可能存在未知的去甲基化酶。因此,新m6A相關酶鑒定仍需要深入的生命科學研究。作為一種重要的RNA表觀遺傳修飾,需要進一步研究m6A如何與DNA甲基化和組蛋白修飾相互作用來調控基因表達,以及m6A修飾與其他RNA修飾之間的潛在聯(lián)系。
本文回顧了家畜經濟性狀中m6A甲基化修飾調控機制的當前知識狀態(tài)。關于這種表觀遺傳機制的現有數據和未來的工作將幫助家畜研究領域的研究人員使用有效方法有效研究RNAm6A修飾水平,從而促進開展調控生物重要功能中關鍵基因表達的研究。利用這些知識,還可以產生新的分子標記或靶點用于定向育種。這些信息可以作為動物經濟性狀遺傳改良的參考,對提高家畜肉類、蛋類和奶類的生產產量和質量具有重要意義。
參考文獻:
Ren T, Xu M, Du X, Wang Y, Loor JJ, Lei L, Gao W, Du X, Song Y, Liu G, Li X. Research Progress on the Role of M6A in Regulating Economic Traits in Livestock. Int J Mol Sci. 2024 Jul 31;25(15) pii: ijms25158365. doi: 10.3390/ijms25158365. PubMed PMID: 39125935.
m6A的基本調控機制
m6A修飾是一種常見的RNA修飾類型,其可以改變RNA分子的化學屬性和空間結構,從而影響其功能性。m6A修飾動態(tài)可逆,參與多種酶之間的復雜互作,包括甲基轉移酶(如METTL3、METTL14和WTAP)、去甲基化酶(如FTO、ALKBH5和ALKBH3)以及m6A結合蛋白(如YTHDF1、YTHDF2、YTHDF3、YTHDC1和YTHDC2)。研究表明,m6A修飾可以影響mRNA的剪接、穩(wěn)定性、出核和翻譯,以及miRNA的加工和功能(圖1) 。
圖1:m6A甲基化修飾的動態(tài)過程。負責添加m6A修飾酶是蛋白質復合體,包括 METTL3、METTL14、WTAP 和其他組分。FTO蛋白是一種去甲基化酶,通過其核心結構域C末端的獨特long loop區(qū)域作用,以去除RNA中的甲基化。ALKBH5是另一種去甲基化酶,通過氧化過程去除帶有m6A修飾的RNA的甲基化。諸如YTHDF1、YTHDF2、YTHDF3、YTHDC1和YTHDC2等reader蛋白參與了包括mRNA剪接、穩(wěn)定性、翻譯和出核在內的各種過程。
m6A 修飾在動物生長發(fā)育中的作用
(1)m6A修飾在肌肉發(fā)育中的調控作用
作為最豐富的mRNA修飾類型,m6A在骨骼肌分化、心肌重塑和再生中發(fā)揮著調控作用。在豬胚胎發(fā)育的6個不同階段中,m6A修飾顯示出高度動態(tài)變化,大多數受影響的基因富集在與骨骼肌發(fā)育相關的通路中。豬產前骨骼肌中的m6A甲基化水平在胚胎年齡E40(原始肌纖維形成期)時顯著降低,隨著胚胎年齡增加(E50-95天,次級肌纖維形成期),甲基化水平逐漸增加。通過siRNA沉默IGF2BP1可以促進增殖并抑制肌細胞分化。骨骼肌發(fā)育在家畜高效肉類生產中的核心地位突顯了m6A修飾對產品質量的重要性。山羊研究表明,FTO可以通過降低m6A修飾程度來增加GADD45B mRNA的穩(wěn)定性和表達,從而激活p38MAPK通路并促進肌細胞分化。METTL3蛋白也可以通過上調m6A水平和增強MEF2C表達來促進山羊骨骼肌衛(wèi)星細胞的肌源性分化。這些研究為通過RNA甲基化調控山羊肌生成提供了潛在的分子機制。
在對牛的研究中,m6A甲基化轉移酶METTL3對成肌細胞增殖具有抑制作用,并能促進細胞分化;它還能通過影響TM4SF1穩(wěn)定性來調控成肌細胞發(fā)育。YTHDF2介導的TM4SF1 mRNA降解也依賴于m6A修飾。成年牛背最長肌中m6A去甲基化酶(FTO和ALKBH5)的表達顯著高于新生牛,而m6A甲基轉移酶(METTL3、METTL14和WTAP)的表達則較低。在肌生成過程中,這五個基因的mRNA表達顯著增加;在牛成肌細胞中的干擾實驗進一步表明,m6A甲基化相關酶可能調控牛骨骼肌發(fā)育。
在京星黃(Jingxing-Huang)雞中,m6A去甲基化酶基因ALKBH5和FTO在骨骼肌發(fā)育過程中的表達水平顯著增加。ALKBH5和FTO蛋白可能通過調控RNA m6A甲基化水平影響雞骨骼肌發(fā)育。有證據表明METTL3、METTL14和WTAP在家禽肌纖維類型組成、維持和成肌細胞分化中具有調控作用。
m6A是影響非編碼RNA(ncRNA)功能的關鍵調控因子,從而參與生物體的生長和發(fā)育。METTL3過表達降低了分化C2C12細胞和肌肉損傷再生小鼠模型中肌肉特異性miR-1a、miR-133a、miR-133b和miR-206表達。研究人員使用RNA-seq和MeRIP-seq技術分析了豬氧化和糖酵解骨骼肌中l(wèi)ncRNAs的m6A甲基化模式。結果顯示,大多數lncRNAs在最后一個外顯子中有一個優(yōu)先富集的m6A peaks,且m6A水平與lncRNAs表達水平之間存在正相關。此外,lncRNA MSTRG.14200.1的抑制延遲了衛(wèi)星細胞分化,并刺激了快速和慢速肌纖維之間的轉換。這項研究為豬肌纖維類型的轉換提供了新的目標。在牛-牦牛骨骼肌中,m6A豐度與基因表達水平呈正相關,但與lncRNA表達水平呈負相關,表明m6A修飾可能在牛-牦牛骨骼肌的發(fā)育中發(fā)揮重要作用,但m6A修飾對mRNA和lncRNA的調控機制可能不同。RNA-seq和ATAC-seq測序技術的使用允許發(fā)現骨骼肌特異性lncRNA MYH1G-AS,其轉錄由轉錄因子SMAD3和SP2協(xié)調。此外,SP2抑制ALKBH5的轉錄,降低了ALKBH5對MYH1G-AS的去甲基化,從而MYH1G-AS RNA穩(wěn)定性失衡。MYH1G-AS加速了成肌細胞的增殖,但抑制其分化;這個基因促進了慢速肌纖維向快速肌纖維的轉換,并導致肌肉萎縮。總體而言,這些一致的結果表明m6A甲基轉移酶在家畜骨骼肌發(fā)育中發(fā)揮重要的調控作用。
(2)m6A修飾在脂肪生成中的調控
有證據表明m6A修飾可以調控轉錄因子和脂肪組織特異性基因的表達水平,從而影響脂肪沉積代謝過程。其中,FTO作為一種RNA去甲基化酶,在修飾基因表達中發(fā)揮重要作用。例如,在豬脂肪細胞中FTO和METTL3基因的過表達和特異性敲除揭示了FTO與m6A甲基化水平呈負相關,而與脂肪生成呈正相關。而METTL3與m6A甲基化水平呈正相關,與脂肪生成呈負相關。在豬骨髓源性間充質干細胞(BMSCs)中,METTL3缺失導致JAK1 mRNA上的m6A修飾減少,進而增強了JAK1 mRNA的穩(wěn)定性和翻譯效率。JAK1-STAT5信號通路是調節(jié)脂肪細胞分化的關鍵通路之一,通過磷酸化STAT5激活與脂肪生成相關的下游基因,如C/EBPβ,并促進其進入細胞核,從而引導BMSCs向脂肪細胞分化。FTO酶通過m6A去甲基化促進豬的前脂肪細胞分化,通過調節(jié)JAK2-STAT3-C/EBPβ信號通路、有絲分裂克隆擴增和細胞自噬通路。在豬肌內前脂肪細胞中,沉默FTO顯著降低了磷酸組蛋白H3水平,并抑制肌內前脂肪細胞增殖。沉默FTO還導致過氧化物酶體增殖物激活受體γ(PPARγ)和CCAAT/增強子結合蛋白α(C/EBPα)下調,以及β-catenin表達的上調。LiCl(一種Wnt/β-catenin信號通路的特異性激活劑)可以減弱FTO誘導的PPARγ上調和β-catenin下調。在豬脂肪細胞中,FTO過表達顯著增加了細胞內甘油三酯含量,并顯著上調了與脂肪生成相關基因的C/EBPβ表達,同時下調了脂肪分解基因ATGL和HSL的mRNA表達;FTO敲低會導致不同結果,表明這種蛋白可能促進豬脂肪細胞中脂質的積累。總之,沉默 FTO 會減少豬肌內脂肪前細胞的增殖和分化,并可能通過 Wnt/β-catenin 通路影響豬肌內脂肪前細胞的分化。
在中國本土雞種中,腹部脂肪過度沉積一直是家禽生產中的一個重大問題。高脂飲食可能導致體重增加和過度脂肪沉積,影響脂肪代謝和脂肪生成的基因和通路。在雞的前脂肪細胞中,RNA m6A甲基化與脂肪生成呈負相關。FTO過表達顯著抑制m6A水平,并促進雞的前脂肪細胞的增殖和分化,而沉默FTO則產生相反結果。FTO酶通過去甲基化調控CTNNB1表達,從而促進雞的脂肪沉積。有研究報告稱,與新生牛相比,秦川成年牛脂肪組織中的m6A甲基轉移酶METTL3、METTL14和WTAP以及m6A去甲基化酶FTO和ALKBH5都顯著上調。它們在體外前脂肪細胞增殖和分化過程中的表達模式也表現出顯著差異。這些結果表明m6A修飾可能在家牛的脂肪沉積中發(fā)揮重要作用,m6A甲基轉移酶在調控牛的前脂肪細胞的增殖和分化中具有潛在作用。
當前研究表明,許多環(huán)狀RNA(circRNA)分子具有多個m6A修飾位點。例如,在黔山黑豬和大白豬的背最長肌中鑒定出12個候選m6A修飾circRNA(m6A-circRNA);這些m6A-circRNA可以作為miRNA海綿分子,并通過構建ceRNA調控網絡來調控脂肪沉積。富集分析顯示,m6A-circRNA的親本基因及其吸附的miRNA靶基因參與與脂肪沉積、細胞增殖和分化相關通路,表明m6A-circRNA在決定豬肉品質方面可能特別重要。這些一致的發(fā)現表明,m6A甲基轉移酶在家牛、豬和家禽的脂肪沉積中發(fā)揮著至關重要的作用。
m6A修飾在家畜繁殖性狀中的功能性作用
(1)m6A修飾對精子發(fā)生的調控
精子發(fā)生是一個高度復雜和特殊的細胞發(fā)育過程,包括精原細胞增殖、精母細胞減數分裂和精子形成三個階段。精子發(fā)生過程中的基因表達在轉錄、轉錄后和翻譯水平上受到嚴格調控,以確保階段特異性基因的準確表達。以往的研究使用m6A RNA甲基轉移酶METTL3和/或METTL14的條件性基因敲除小鼠模型揭示了METTL3或METTL14在早期精原細胞中的缺失會導致精原干細胞凋亡。然而,只有在相對較晚階段的精原細胞(A1型精原細胞)中同時敲除METTL3和METTL14,才會影響精子形成。在人類中,WTAP和ALKBH5表達在精子發(fā)生過程中減少;FTO和ALKBH5在哺乳動物的睪丸中高表達,人類FTO基因突變與精液質量下降密切相關。有研究報告稱,ALKBH5基因缺陷的雄性小鼠由于減數分裂中期精原細胞凋亡而生育能力降低。山羊睪丸發(fā)育中ALKBH5表達上調,山羊精原干細胞中ALKBH5過表達促進了從G1期到S期轉變并抑制凋亡。研究人員使用m6A-seq、RNA-Seq和核糖體分析技術,生成了精子發(fā)生不同發(fā)育階段m6A RNA修飾的動態(tài)圖譜。研究結果表明m6A甲基轉移酶失活導致m6A RNA修飾水平下降,從而導致參與精原細胞命運決定和精子形成的基因轉錄和翻譯效率發(fā)生顯著變化。
轉錄延伸標記H3K36me3介導m6A修飾的共轉錄過程。H3K36me3與m6A甲基轉移酶復合體中的METTL3和METTL14互作,促進新生RNA m6A修飾。組蛋白和RNA修飾之間的這種交流為mRNA-m6A的生物發(fā)生提供了新的概念。盡管DNA甲基化和m6A修飾是獨立過程,但它們可能共同影響基因表達和精子發(fā)生的調控。非編碼RNA作為表觀遺傳調控因子,在精子發(fā)生過程中起著重要作用。其中,miRNA、circRNA和PiRNA在轉錄、轉錄后和翻譯水平上調節(jié)雄性生殖細胞的正常發(fā)育,并可能與m6A修飾互作。先前的研究表明,在小鼠GC-1精原細胞中,敲低METTL3后,構建的circRNA-miRNA-mRNA調控網絡揭示了H2afx和Dnmt3a是精子減數分裂中的樞紐基因。這項研究可能為由異常m6A修飾引起的異常精子發(fā)生的病理機制提供新的見解和潛在的治療靶點。
豬生殖細胞中的所有mRNA轉錄本都含有m6A修飾,m6A的reads密度在編碼序列(CDS)中增加,并在終止密碼子附近達到最高值,然后在3'非翻譯區(qū)(UTR)中降低。在精子發(fā)生過程中,轉錄產物中m6A修飾的豐度隨著不同發(fā)育階段而變化。粗線期精母細胞和圓形精子細胞中CDS的m6A修飾reads密度較高,而在精原細胞的30非編碼區(qū)中較高。METTL3缺失會導致精原細胞分化異常啟動,并影響精母細胞在減數分裂前達到粗線期的能力。在粗線期精母細胞中,甲基化基因上調優(yōu)先參與精子發(fā)生和細胞周期過程,這些過程被m6A修飾上調,表明m6A在早期發(fā)育階段的重要作用。此外,PS中的m6A修在參與精子細胞發(fā)育基因中富集,并上調參與微管加工、纖毛組織和纖毛組裝的基因表達,表明m6A甲基化mRNA可能在精子發(fā)生前抑制翻譯。在圓形精子細胞中,m6A甲基化基因上調,并調控包括多細胞生物解剖結構和管狀形態(tài)發(fā)生在內的發(fā)育過程,表明m6A修飾在介導豬精子發(fā)生中的保守性。在雄性生殖細胞發(fā)育過程中的m6A RNA修飾保守并受動態(tài)調控,廣泛存在于精子發(fā)生的各個階段,特別是在粗線期/雙線期精母細胞和圓形精子細胞中高度富集。在牛、牦牛和牦牛-牛雜交品種的睪丸組織中,bta-miR-200a通過抑制ALKBH5的表達調控m6A甲基化水平,從而影響m6A靶基因的表達和功能。這些一致的結果表明m6A甲基轉移酶可能參與動物精子發(fā)生(圖3)。
(2)m6A修飾對動物卵母細胞成熟的調控
動物卵母細胞成熟可以分為三個主要階段:減數分裂成熟、細胞質成熟和表觀遺傳成熟。卵母細胞的表觀遺傳成熟主要包括調控基因表達的DNA甲基化和組蛋白修飾。m6A甲基化修飾可以影響mRNA降解速率和轉錄本長度,并阻礙正常的mRNA翻譯。這可能導致卵泡發(fā)育缺陷、卵母細胞成熟抑制、卵母細胞早期發(fā)育停滯和合子基因組激活受損,從而影響和調控雌性哺乳動物的卵母細胞形成各個階段。
卵母細胞支持胚胎發(fā)育能力在卵泡生成過程中獲得,與卵泡發(fā)育過程中周圍顆粒細胞的互作可以影響卵母細胞質量。豬顆粒細胞表現出豐富的m6A基因修飾,從小卵泡(<3毫米)到大卵泡(>5毫米)的轉變過程中,m6A甲基化發(fā)生動態(tài)變化。在小卵泡和大卵泡的顆粒細胞中分別鑒定出7289和6882個m6A位點,m6A在5'UTR或3'UTR附近富集并共有保守motif。進一步分析顯示,差異表達的m6A甲基化基因在與類固醇生成、顆粒細胞增殖和卵泡發(fā)育相關的多個信號通路中顯著富集。總體而言,這些結果突出了豬卵泡發(fā)育過程中顆粒細胞中存在不同的m6A修飾,這可能與類固醇和卵泡生成有關。
牦牛的發(fā)情周期是一種季節(jié)性現象,可能涉及性激素合成和分泌以及對卵泡生長和發(fā)育的表觀遺傳調控。HE染色顯示,在發(fā)情期卵巢中的發(fā)育中卵泡和成熟卵泡數量明顯多于黃體期和妊娠期。qPCR結果顯示,在卵巢發(fā)育的不同階段,METTL3、METTL14、FTO和YTHDC1的表達存在顯著差異,表明m6A修飾在卵巢活動中具有調控作用。通過MERIP-seq技術繪制的牦牛m6A轉錄組譜顯示,在編碼序列(CDS)、3'UTR和保守序列“DRACH”中m6A peaks高度富集。GO、KEGG和GSEA分析結果表明,m6A修飾可能參與牦牛卵巢在生殖周期中的許多生理活動。
在家禽研究中,卵泡內的體細胞決定了卵泡的選擇過程。在雞的卵泡選擇過程中對m6A甲基化水平分析顯示,顆粒細胞和卵泡膜細胞中mRNA甲基化和去甲基化廣泛存在,m6A甲基化修飾與基因表達水平呈負相關。在卵泡選擇過程中,m6A甲基化peaks和修飾轉錄本均增加,導致與卵泡生成相關的許多基因表達動態(tài)變化。功能富集分析表明,Wnt通路中關鍵因子的m6A修飾可能在雞的卵泡選擇中發(fā)揮主要作用。
先前研究表明,用m6A抑制劑(環(huán)亮氨酸)處理卵母細胞可以抑制RNA m6A甲基化和組蛋白修飾,并改變囊胚階段代謝基因表達,從而抑制豬卵母細胞的減數分裂和早期胚胎發(fā)育。這些一致的結果表明,m6A甲基轉移酶可能參與主要家畜物種卵母細胞的成熟過程。
總結和觀點
m6A修飾,作為一種廣泛存在的重要表觀遺傳修飾,在農業(yè)動物的各種生命過程中扮演著重要角色,包括生長和發(fā)育、脂質代謝和繁殖(表1)。
表1:家畜中主要的m6A甲基化相關酶。
近年來,多組學整合分析已成為生命科學領域最重要的研究方法之一。越來越多的研究關注m6A修飾與其他組學數據的綜合分析。例如,轉錄組與m6A之間的相關性可以全面解釋轉錄因子對基因表達和表型差異的調控作用。在子宮內膜癌研究中,通過MeRIP-seq、RNA-seq測序和實驗分析,發(fā)現IGF2BP1鑒定目標基因PEG10終止密碼子附近的m6A甲基化位點,從而調控子宮內膜癌細胞增殖。此外利用全基因組、RNA-seq和功能研究,揭示IGF2BP1基因啟動子區(qū)域缺失可能是與雞體型相關數量性狀位點的因果變異。
與人類和小鼠的m6A修飾研究相比,家畜的m6A修飾研究相對較少。目前,使用m6A-seq評估m(xù)6A修飾在家畜經濟性狀中的作用的研究主要集中在甲基轉移酶、m6A水平以及與性狀相關的差異甲基化基因鑒定上。因此,需要進一步研究家畜中RNA甲基化相關酶和甲基化基因的具體功能和調控機制。RNA甲基化研究仍面臨許多挑戰(zhàn),如m6A-seq測序技術限制可能導致的假陽性結果,以及缺乏對家畜RNA甲基化動態(tài)性質和調控機制的研究。
m6A修飾通常被視為基因表達的調控因子和許多調控通路中的目標。m6A的writers、erasers、readers經常相互交互。在癌癥研究中,METTL3可能調控WTAP蛋白穩(wěn)定性,而WTAP蛋白上調只有在METTL3存在時才具有致癌作用。參與不同類型癌癥的m6A相關蛋白通常受到多種m6A蛋白因子調控。由于對m6A了解有限,仍需要廣泛研究通路之間的互作以及m6A在調控基因表達中的協(xié)同效應。迄今為止,已經發(fā)現了多種writers,但只鑒定出兩種erasers:FTO和ALKBH5。FTO或ALKBH5的異常表達只會導致生物體整體m6A修飾水平的微小變化,這表明體內可能存在未知的去甲基化酶。因此,新m6A相關酶鑒定仍需要深入的生命科學研究。作為一種重要的RNA表觀遺傳修飾,需要進一步研究m6A如何與DNA甲基化和組蛋白修飾相互作用來調控基因表達,以及m6A修飾與其他RNA修飾之間的潛在聯(lián)系。
本文回顧了家畜經濟性狀中m6A甲基化修飾調控機制的當前知識狀態(tài)。關于這種表觀遺傳機制的現有數據和未來的工作將幫助家畜研究領域的研究人員使用有效方法有效研究RNAm6A修飾水平,從而促進開展調控生物重要功能中關鍵基因表達的研究。利用這些知識,還可以產生新的分子標記或靶點用于定向育種。這些信息可以作為動物經濟性狀遺傳改良的參考,對提高家畜肉類、蛋類和奶類的生產產量和質量具有重要意義。
參考文獻:
Ren T, Xu M, Du X, Wang Y, Loor JJ, Lei L, Gao W, Du X, Song Y, Liu G, Li X. Research Progress on the Role of M6A in Regulating Economic Traits in Livestock. Int J Mol Sci. 2024 Jul 31;25(15) pii: ijms25158365. doi: 10.3390/ijms25158365. PubMed PMID: 39125935.
標簽:
RNA甲基化



Copyright(C) 1998-2025 生物器材網 電話:021-64166852;13621656896 E-mail:info@bio-equip.com