
®”(dЁЎng)З°О»ЦГ > КЧн“ > јјРg(shЁҙ)ОДХВ > RNA-BSФЪҪТКҫDNMT1ФЪm5CРЮп—ЦРөДҫҖБЈуw№ҰДЬҷCЦЖЦРөД‘Ә(yЁ©ng)УГ
RNA-BSФЪҪТКҫDNMT1ФЪm5CРЮп—ЦРөДҫҖБЈуw№ҰДЬҷCЦЖЦРөД‘Ә(yЁ©ng)УГ
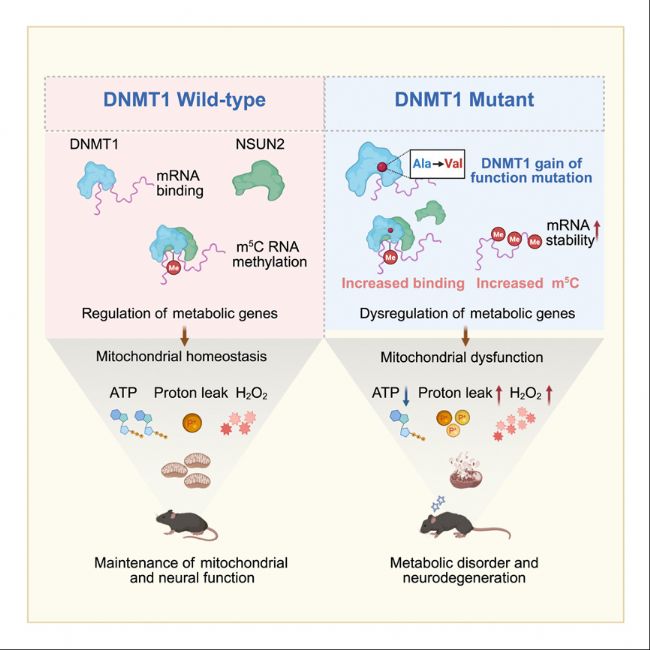
ұнУ^ЯzӮчХ{(diЁӨo)ҝШЈ¬°ьАЁDNAјЧ»щ»ҜЎўRNAРЮп—әНИҫЙ«Щ|(zhЁ¬)ЦШЛЬөИЈ¬ФЪЙсҪӣ(jЁ©ng)НЛРРРФјІІЎЦРЖрЦШТӘЧчУГЎЈDNAјЧ»щЮD(zhuЁЈn)ТЖГё1ЈЁDNMT1Ј©КЗТ»·NТСЦӘөДDNAјЧ»щЮD(zhuЁЈn)ТЖГёЈ¬ЦчТӘШ“Шҹ(zЁҰ)ҫSіЦDNAјЧ»щ»ҜЎЈИ»¶шDNMT1ФЪ·З·ЦБСјҡ°ыЦРөД№ҰДЬЙРІ»ЗеіюЎЈҙЛНвЈ¬DNMT1өДRFTSЈЁreplication focus targeting sequence Ј©ҪY(jiЁҰ)ҳӢ(gЁ°u)УтН»ЧғЈЁИзA560VЈ©Еc¶а·NЙсҪӣ(jЁ©ng)НЛРРРФјІІЎПакP(guЁЎn)Ј¬ө«Жд·ЦЧУҷCЦЖЙРОҙНкИ«кUГчЎЈ
ҪьИХЈ¬ГАҮшјУЦЭҙуҢW(xuЁҰ)ВеЙјҙү·ЦРЈҪKЙнҪМКЪ/ЙПәЈҝЖјјҙуҢW(xuЁҰ)ГвТЯ»ҜҢW(xuЁҰ)СРҫҝЛщМШЖёҪМКЪ·¶ҮшЖҪХnо}ҪMҪТКҫБЛ DNMT1ФЪХ{(diЁӨo)ҝШDNAәНRNAјЧ»щ»ҜЦРөДлpЦШЧчУГЈ¬МШ„eКЗЖдНЁЯ^RNAРЮп—Х{(diЁӨo)ҝШҫҖБЈуw№ҰДЬөДҷCЦЖЎЈСРҫҝ°l(fЁЎ)¬F(xiЁӨn)Ј¬DNMT1ДЬүтЕcmRNAҪY(jiЁҰ)әПІўҙЩЯMЖд·Җ(wЁ§n)¶ЁРФЈ¬ІўНЁЯ^ХРДјNSUN2ө°°ЧҒнХ{(diЁӨo)№қ(jiЁҰ)RNAөД5-јЧ»щ°ыаЧаӨЈЁm5CЈ©јЧ»щ»ҜЎЈҙЛНвЈ¬СРҫҝЯҖ°l(fЁЎ)¬F(xiЁӨn)DNMT1өДRFTSҪY(jiЁҰ)ҳӢ(gЁ°u)УтН»ЧғЈЁИзA560VЈ©•юҢ§(dЁЈo)ЦВҙъЦx»щТтөДRNAјЧ»щ»ҜәН·Җ(wЁ§n)¶ЁРФ®җіЈЈ¬ЯM¶шТэ°l(fЁЎ)ҫҖБЈуw№ҰДЬХПөKәНЙсҪӣ(jЁ©ng)НЛРРРФјІІЎЎЈЯ@Р©°l(fЁЎ)¬F(xiЁӨn)һйАнҪвDNMT1ФЪЙсҪӣ(jЁ©ng)ПөҪy(tЁҜng)јІІЎЦРөДЧчУГМṩБЛРВөДТ•ҪЗЈ¬Іўһйй_°l(fЁЎ)°РПтRNAјЧ»щ»ҜөДЦОҜҹІЯВФМṩБЛАнХ“»щөA(chЁі)ЎЈПакP(guЁЎn)СРҫҝіЙ№ыТФЎ¶DNA methyltransferase 1 modulates mitochondrial function through bridging m5C RNA methylationЎ·һйо}°l(fЁЎ)ұнУЪЎ¶Molecular CellЎ·ЖЪҝҜЎЈ

ОДХВҳЛ(biЁЎo)о}ЈәDNA methyltransferase 1 modulates mitochondrial function through bridging m5C RNA methylation
У°н‘ТтЧУЈәIF14.5/Q1
јјРg(shЁҙ)ЖҪЕ_Јә RNA-BS-seqЎўRRBSЎўRNA-seqөИ

DNMT1КЗТ»·NҫSіЦDNAјЧ»щ»ҜөДDNAјЧ»щЮD(zhuЁЈn)ТЖГёЎЈЖдҸН(fЁҙ)ЦЖҪ№ьc°РПтРтБРЈЁRFTSЈ©ҪY(jiЁҰ)ҳӢ(gЁ°u)УтЦРөДьcН»Чғ•юҢ§(dЁЈo)ЦВНн°l(fЁЎ)РФЙсҪӣ(jЁ©ng)НЛРРРФјІІЎЈ¬ИзіЈИҫЙ«уwп@РФРЎДXРФ№ІқъК§Х{(diЁӨo)-¶ъГ@әНКИЛҜ°YЈЁADCA-DNЈ©јІІЎЎЈұҫСРҫҝКЧПИтһЧCБЛDNMT1ҫЯУРҪY(jiЁҰ)әПmRNAЮD(zhuЁЈn)дӣұҫөД№ҰДЬЈ¬ІўНЁЯ^ХРДјNOP2/Sun RNAјЧ»щЮD(zhuЁЈn)ТЖГё2ЈЁNSUN2Ј©ҒнҙЩЯMRNA m5CјЧ»щ»ҜЎЈН¬•rЈ¬RNA m5CјЧ»щ»ҜФцҸҠДЗР©Х{(diЁӨo)№қ(jiЁҰ)ҫҖБЈуw№ҰДЬ»щТтөДRNA·Җ(wЁ§n)¶ЁРФЎЈ®”(dЁЎng)РЎКуөДDNMT1 RFTSҪY(jiЁҰ)ҳӢ(gЁ°u)Ут°l(fЁЎ)ЙъН»Чғ•rЈ¬•юТэ°l(fЁЎ)®җіЈөДDNMT1-RNAПа»ҘЧчУГЈ¬Іўп@ЦшМбёЯІҝ·ЦҙъЦx»щТтөДm5C RNAјЧ»щ»ҜәНRNA·Җ(wЁ§n)¶ЁРФЎЈТтҙЛЈ¬ҙъЦxRNAЮD(zhuЁЈn)дӣұҫЛ®ЖҪФцјУҢ§(dЁЈo)ЦВБЛАЫ·eРФөДСх»Ҝ‘Ә(yЁ©ng)јӨЎўҫҖБЈуw№ҰДЬХПөKәНЙсҪӣ(jЁ©ng)°Y оЎЈҝӮуw¶шСФЈ¬ұҫСРҫҝҪY(jiЁҰ)№ыҪТКҫБЛDNMT1ФЪХ{(diЁӨo)№қ(jiЁҰ)DNAәНRNAјЧ»щ»ҜЦРөДлpЦШЧчУГЈ¬ІўЯMТ»ІҪХ{(diЁӨo)№қ(jiЁҰ)БЛҫҖБЈуw№ҰДЬЈ¬һйDNMT1Н»ЧғХTҢ§(dЁЈo)өДЙсҪӣ(jЁ©ng)НЛРРРФјІІЎөД°l(fЁЎ)ІЎҷCЦЖМṩБЛРВТҠҪвЎЈ
СРҫҝ·Ҫ·Ё
1. јҡ°ыЕарB(yЁЈng)әН»щТтҫҺЭӢ
јҡ°ыЕарB(yЁЈng)ЈәК№УГHeLaјҡ°ыЎўHEK293Tјҡ°ыТФј°РЎКуЕЯМҘёЙјҡ°ыЈЁESCsЈ©Ј¬ІўФЪМШ¶Ё—lјюПВЕарB(yЁЈng)ЎЈ
»щТтҫҺЭӢЈәНЁЯ^CRISPR-Cas9јјРg(shЁҙ)ҳӢ(gЁ°u)ҪЁБЛ”yҺ§Dnmt1A560VН»ЧғөДРЎКуДЈРНәНјҡ°ыПөЎЈ
2Ўў„УОпДЈРНәНРРһйҢW(xuЁҰ)ңyФҮ
РЎКуДЈРНЈәҳӢ(gЁ°u)ҪЁDnmt1A560VН»ЧғРЎКуДЈРНЈ¬УГУЪСРҫҝDNMT1Н»ЧғҢҰЙсҪӣ(jЁ©ng)ПөҪy(tЁҜng)өДйLЖЪУ°н‘ЎЈ
РРһйҢW(xuЁҰ)ңyФҮЈә°ьАЁәуЦ«ЧҘОХңyФҮәНРэЮD(zhuЁЈn)—UңyФҮЈ¬Фu№АРЎКуөДЯ\„У№ҰДЬХПөKЎЈ
3. RNAҪY(jiЁҰ)әПө°°Ч·ЦОц
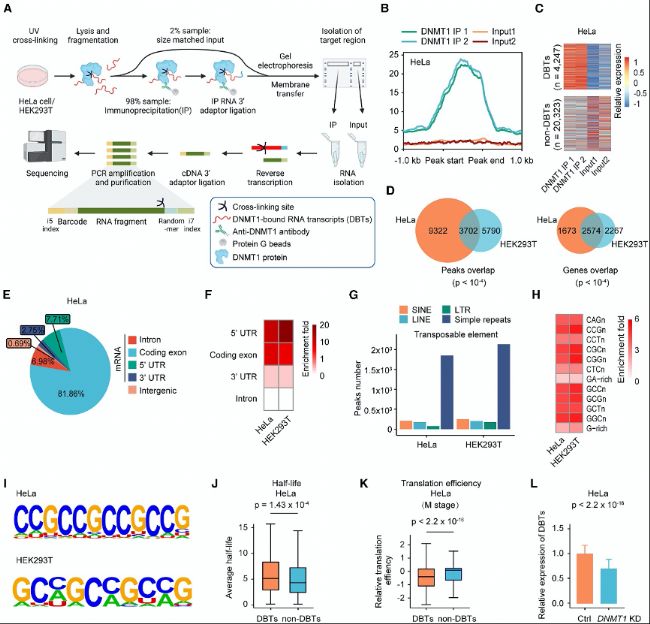
ФцҸҠРНҪ»В“(liЁўn)ГвТЯіБөнңyРтЈЁeCLIP-seqЈ©Јәиb¶ЁDNMT1ҪY(jiЁҰ)әПөДmRNAЮD(zhuЁЈn)дӣұҫЈЁDNMT1-bound mRNA transcriptsЈ¬DBTsЈ©Ј¬°l(fЁЎ)¬F(xiЁӨn)DNMT1ДЬүтҪY(jiЁҰ)әПҙуБҝmRNAЈ¬ІўУ°н‘Жд·Җ(wЁ§n)¶ЁРФЎЈ
4. RNAјЧ»щ»Ҝ·ЦОц
RNA-BS-seqЈә·ЦОцRNA m5CјЧ»щ»ҜЛ®ЖҪЈ¬°l(fЁЎ)¬F(xiЁӨn)DNMT1ДЬүтНЁЯ^ХРДјNSUN2ө°°ЧҒнХ{(diЁӨo)№қ(jiЁҰ)RNAөДm5CјЧ»щ»ҜЎЈ
Щ|(zhЁ¬)ЧV·ЦОцЈЁUHPLC-MRM-MS/MSЈ©Јә¶ЁБҝ·ЦОцRNAјЧ»щ»ҜЛ®ЖҪЈ¬тһЧCDNMT1ҢҰm5CјЧ»щ»ҜөДХ{(diЁӨo)ҝШЧчУГЎЈ
5. ө°°ЧЩ|(zhЁ¬)»ҘЧч·ЦОц
Co-IPЈәҷzңyDNMT1ЕcNSUN2»ҘЧчЈ¬ІўНЁЯ^Щ|(zhЁ¬)ЧV·ЦОциb¶ЁDNMT1өД»ҘЧчө°°ЧЎЈ
өИңШөО¶ЁБҝҹб·ЁЈЁITCЈ©ЈәҷzңyDNMT1ЕcNSUN2Ц®йgөДҪY(jiЁҰ)әПУHәНБҰЎЈ
6. »щТтұнЯ_әНұнУ^ЯzӮч·ЦОц
RNA-seqЈә·ЦОц»щТтұнЯ_Чғ»ҜЈ¬°l(fЁЎ)¬F(xiЁӨn)Dnmt1A560VН»ЧғҢ§(dЁЈo)ЦВҙъЦx»щТтұнЯ_К§Х{(diЁӨo)ЎЈ
DNAјЧ»щ»ҜңyРтЈЁEM-seqәНRRBSЈ©Јә·ЦОцDNAјЧ»щ»ҜЛ®ЖҪЈ¬°l(fЁЎ)¬F(xiЁӨn)Н»ЧғҢҰDNAјЧ»щ»ҜУ°н‘Э^РЎЈ¬ө«ҢҰRNAјЧ»щ»ҜУ°н‘п@ЦшЎЈ
7. ҶОјҡ°ы·ЦОц
scRNA-seqәНsnRNA-seqЈә·ЦОцЙсҪӣ(jЁ©ng)ҪMҝ—ЦРІ»Н¬јҡ°ыоҗРНөД»щТтұнЯ_Чғ»ҜЈ¬ҪТКҫБЛҸV·әөДСх»Ҝ‘Ә(yЁ©ng)јӨ·ҙ‘Ә(yЁ©ng)ЎЈ
8. ҙъЦxәНҫҖБЈуw№ҰДЬ·ЦОц
јҡ°ыДЬБҝҙъЦx·ЦОцЈәФu№Ајҡ°ыөДСхҡвПыәДВКЈЁOCRЈ©әН°ыНвЛб»ҜВКЈЁECARЈ©ЎЈ
ҫҖБЈуwDNAә¬Бҝңy¶ЁЈәНЁЯ^qPCR·ЦОцҫҖБЈуwDNAә¬БҝЎЈ
Сх»Ҝ‘Ә(yЁ©ng)јӨәНATPә¬Бҝ·ЦОцЈәҷzңyјҡ°ыЦРөДСх»Ҝ‘Ә(yЁ©ng)јӨәНATPЛ®ЖҪЎЈ
ҪY(jiЁҰ)№ыҲDРО
ЈЁ1Ј©DNMT1ЦұҪУЕcmRNAҪY(jiЁҰ)әПТФФцҸҠЖд·Җ(wЁ§n)¶ЁРФ
DNMT1ДЬүтЕcmRNAҪY(jiЁҰ)әПЈ¬ІўНЁЯ^ФцҸҠRNA·Җ(wЁ§n)¶ЁРФТФХ{(diЁӨo)ҝШ»щТтұнЯ_ЎЈНЁЯ^eCLIP-seqјјРg(shЁҙ)Ј¬СРҫҝХЯФЪHeLaјҡ°ыәНHEK293Tјҡ°ыЦРиb¶ЁіцҙуБҝDBTsЈ¬Я@Р©ЮD(zhuЁЈn)дӣұҫЦчТӘ·ЦІјФЪ5’UTR…^(qЁұ)УтЎЈDNMT1өДҪY(jiЁҰ)әПп@ЦшФцҸҠБЛЯ@Р©mRNAөД·Җ(wЁ§n)¶ЁРФЈ¬ЗТЯ@·N·Җ(wЁ§n)¶ЁРФЕcRNAөДјҡ°ыәЛЩ|(zhЁ¬)ұИәН·ӯЧgР§ВКПакP(guЁЎn)ЎЈ
ЈЁ2Ј©DNMT1НЁЯ^ХРДјNSUN2Х{(diЁӨo)ҝШDBTsөДm5C RNAјЧ»щ»Ҝ
DNMT1ДЬүтНЁЯ^ХРДјNSUN2ө°°ЧҒнХ{(diЁӨo)№қ(jiЁҰ)RNA m5CјЧ»щ»ҜЎЈНЁЯ^ГвТЯ№ІіБөнҢҚтһәНЩ|(zhЁ¬)ЧV·ЦОцЈ¬СРҫҝХЯ°l(fЁЎ)¬F(xiЁӨn)DNMT1ЕcNSUN2Ц®йgҙжФЪЦұҪУПа»ҘЧчУГЈ¬ЗТЯ@·NПа»ҘЧчУГІҝ·ЦТАЩҮУЪRNAЎЈRNA-BS-seq·ЦОцп@КҫЈ¬DNMT1ЗГөНЈЁKDЈ©өДјҡ°ыЦРm5CЛ®ЖҪп@ЦшҪөөНЈ¬ЗТЯ@Р©О»ьcЕcDBTsёЯ¶ИЦШҜBЎЈ
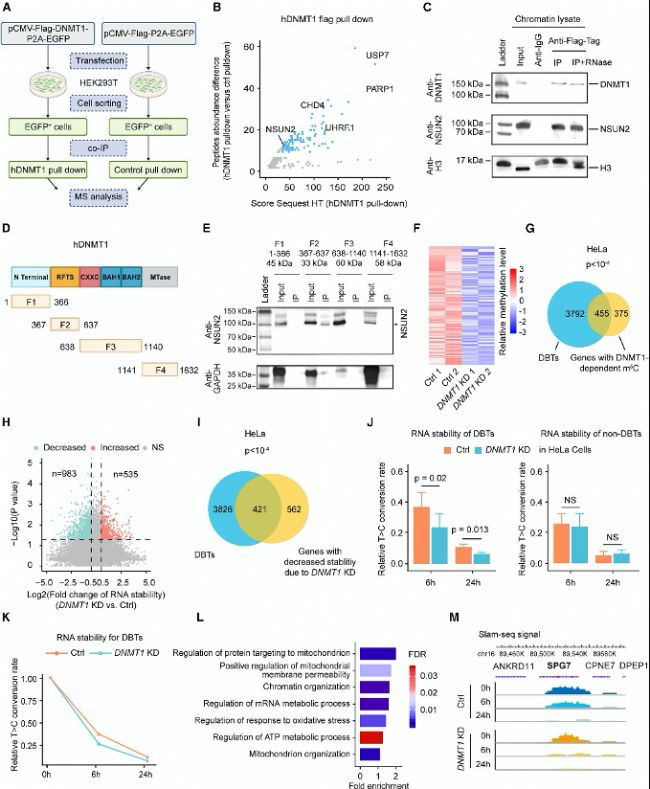
(B) ФЪIP-MSҢҚтһЦРЈ¬НЁЯ^Щ|(zhЁ¬)ЧVҷzңyөҪөДDNMT1Па»ҘЧчУГлД¶ОЕc·ЗМШ®җРФлД¶ОЈЁғHEGFPЈ©өДШS¶ИІо®җЎЈ
(C) ФЪHeLaјҡ°ыЦР®җО»ұнЯ_И«йLNSUN2әНFLAGҳЛ(biЁЎo)әһөДDNMT1ЎЈлSәуК№УГҝ№FLAGҝ№уwЯMРРИҫЙ«Щ|(zhЁ¬)БСҪвТәөДIPЎЈИ»әуУГҝ№NSUN2әНҝ№DNMT1ҝ№уwҢҰIPө°°ЧЯMРРГвТЯҷzңyЎЈ
(D) DNMT1ҪY(jiЁҰ)ҳӢ(gЁ°u)УтәНFLAG-DNMT1·ЦлxЖ¬¶ОЈЁF1–F4Ј©өДКҫТвҲDЎЈ
(E) №ІГвТЯіБөнҷzңyFLAG-DNMT1·ЦлxЖ¬¶ОЈЁF1–F4Ј©ЕcNSUN2өДПа»ҘЧчУГЎЈ
(F) RNA-BS-seqҹбҲDп@КҫФЪHeLaјҡ°ыЦРиb¶ЁіцөДТАЩҮDNMT1өДm5CО»ьcЎЈ
(G) DBTsЕcә¬УРDNMT1ТАЩҮРФm5CО»ьcөД»щТтЦ®йgөДЦШҜBЎЈ
(H) УЙУЪDNMT1ЗГөНЈЁKDЈ©МҺАн¶шФЪHeLaјҡ°ыЦР·Җ(wЁ§n)¶ЁРФ°l(fЁЎ)ЙъЧғ»ҜөДЮD(zhuЁЈn)дӣұҫөДиb¶ЁЎЈ
(I) DBTsЕcТтDNMT1 KD¶шRNA·Җ(wЁ§n)¶ЁРФҪөөНөД»щТтЦ®йgөДЦШҜBЎЈ
(J) ҢҰХХҪMЈЁCtrlЈ©әНDNMT1 KD HeLaјҡ°ыЦРDBTsЈЁЧуӮИ(cЁЁ)Ј©әН·ЗDBTsЈЁУТӮИ(cЁЁ)Ј©өДRNA·Җ(wЁ§n)¶ЁРФ—lРОҲDЎЈ
(K) ҢҰХХҪMЈЁCtrlЈ©әНDNMT1 KD HeLaјҡ°ыЦРЛщУРDBTsөДRNA·Җ(wЁ§n)¶ЁРФҝӮҪY(jiЁҰ)ЎЈ
(L) ҢҰә¬УРDNMT1ТАЩҮРФm5CО»ьcөДDBTsЯMРРGO·ЦОцЎЈ
(M) »щТтҪMЬүЫEп@КҫФЪҢҰХХҪMәНDNMT1 KD HeLaјҡ°ыЦРSlam-seqРЕМ–ФЪSPG7ЙПөД·ЦІјЎЈ
ЈЁ3Ј©Dnmt1A560VН»ЧғРЎКуіц¬F(xiЁӨn)ҙъЦxәНЙсҪӣ(jЁ©ng)ПөҪy(tЁҜng)јІІЎ
Dnmt1A560VН»ЧғРЎКуұн¬F(xiЁӨn)іц¶а·NҙъЦxәНЙсҪӣ(jЁ©ng)°Y оЈ¬°ьАЁуwЦШПВҪөЎўЯ\„У№ҰДЬХПөKәНСх»Ҝ‘Ә(yЁ©ng)јӨФцјУЎЈЯ@Р©°Y оЕcҫҖБЈуw№ҰДЬХПөKПакP(guЁЎn)Ј¬ЗТФЪН»ЧғРЎКуөД¶аӮҖҪMҝ—ЦРУ^ІмөҪҙъЦx»щТтөДRNAјЧ»щ»ҜәН·Җ(wЁ§n)¶ЁРФп@ЦшФцјУЎЈ
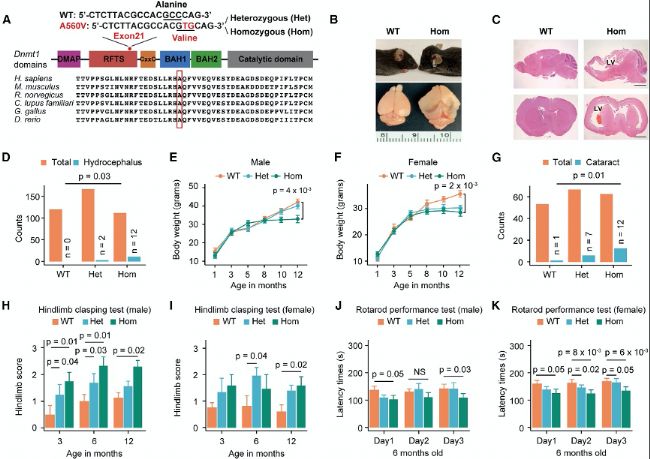
(B) Іҝ·ЦјғәПН»ЧғЈЁHomЈ©РЎКуұн¬F(xiЁӨn)іцҲAн” оо^пBөДДX·eЛ®ЈЁЙПЈ©ЎЈХыуwҙуДXп@Кҫіц”UҙуөДҙуДX°лЗтЎўКЬүәөДРбЗтәНПВПЭөДҙуДXЖӨҢУЈЁПВЈ©ЎЈ
(C) ҙъұнРФҙуДXКё оГжЈЁЙПЈ©әН№Ъ оГжЈЁПВЈ©ЗРЖ¬п@КҫЈ¬ЕcТ°ЙъРНЈЁWTЈ©РЎКуПаұИЈ¬HomН»ЧғРЎКуөДӮИ(cЁЁ)ДXКТЈЁLVsЈ©ҳO¶И”UҸҲЈ¬ЗТҙуДXЖӨҢУЧғұЎЎЈ
(D) ұҫСРҫҝЦРиb¶ЁіцөДWTЎўHetәНHomРЎКуөДҝӮ”ө(shЁҙ)әНДX·eЛ®РЎКуөД”ө(shЁҙ)БҝЎЈ
(E-F) РЫРФРЎКуЈЁГҝ·N»щТтРНn=20Ј©ЈЁEЈ©әНҙЕРФРЎКуЈЁГҝ·N»щТтРНn=20Ј©ЈЁFЈ©өДЙъйLЗъҫҖЎЈ
(G) ұҫСРҫҝЦРИэ·N»щТтРНөД°ЧғИ(nЁЁi)ХП°l(fЁЎ)ІЎВКЎЈ
(H-I) Иэ·N»щТтРНРЎКуөДәуЦ«ЧҘОХФu·ЦЈ¬РЫРФЈЁHЈ©әНҙЖРФЈЁIЈ©ЎЈ
(J-K) 6ӮҖФВҙуРЫРФЈЁJЈ©әНҙЖРФЈЁKЈ©РЎКуөДРэЮD(zhuЁЈn)—UРФДЬңyФҮЎЈ
ЈЁ4Ј©Dnmt1A560VН»Чғп@ЦшФцјУҙъЦxmRNAЮD(zhuЁЈn)дӣұҫЛ®ЖҪ
ФЪDnmt1A560VН»ЧғРЎКујҡ°ыЦРЈ¬ҙъЦx»щТтөДmRNAЮD(zhuЁЈn)дӣұҫЛ®ЖҪп@ЦшФцјУЎЈЯ@Р©ЮD(zhuЁЈn)дӣұҫөДФцјУЕcRNA m5CјЧ»щ»ҜЛ®ЖҪЙэёЯПакP(guЁЎn)Ј¬ұнГчDNMT1Н»ЧғФцҸҠБЛRNAјЧ»щ»ҜәН·Җ(wЁ§n)¶ЁРФЎЈ
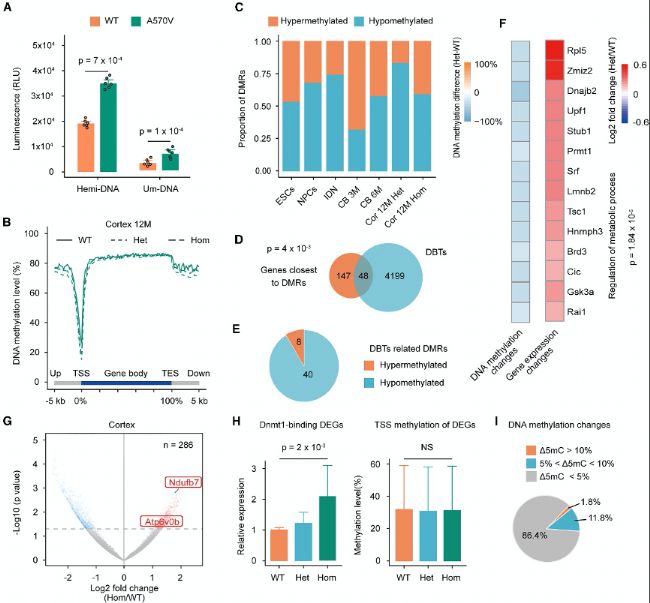
(B) І»Н¬РЎКуҪMҝ—ЦР»щТтуwөДЖҪҫщјЧ»щ»ҜЛ®ЖҪЎЈCBЈ¬РЎДXЈ»CorЈ¬ЖӨҢУЈ»MЈ¬ФВэgЎЈ
(C) І»Н¬ҳУұҫЦРёЯјЧ»щ»ҜәНөНјЧ»щ»ҜDMRsұИАэЎЈ
(D) ЧоҪУҪьDMRs»щТтЕcDBTsөДЦШҜBЎЈ
(E) ЖӨҢУЦРЕcDBTПакP(guЁЎn)өДDMRs”ө(shЁҙ)БҝөДпһҲDЎЈ
(F) ЖӨҢУЦРЕcDBTПакP(guЁЎn)өДDMRsөДјЧ»щ»ҜІо®җЈЁЧуЈ©әНПа‘Ә(yЁ©ng)өД»щТтұнЯ_Чғ»Ҝұ¶”ө(shЁҙ)ЈЁУТЈ©ЎЈ
(G) иb¶ЁЖӨҢУЦРөДІо®җұнЯ_»щТтЈЁDEGsЈ©ЎЈ
(H) ЛщУРЕcDBTПакP(guЁЎn)өДDEGsөДПаҢҰұнЯ_ЈЁЧуЈ©әНЖӨҢУЦРПа‘Ә(yЁ©ng)TSS±2 kb…^(qЁұ)УтөДјЧ»щ»ҜЛ®ЖҪЈЁУТЈ©ЎЈ
(I) ҢҰЖӨҢУЦРЕcDBTПакP(guЁЎn)өДDEGsөДTSS±2kb…^(qЁұ)УтЦРЈ¬јғәПЧУЈЁHomЈ©ЕcТ°ЙъРНЈЁWTЈ©Ц®йgөДDNAјЧ»щ»ҜЧғ»ҜұИАэ·ЦОцЎЈ
ЈЁ5Ј©·Җ(wЁ§n)¶Ё»ҜөДDNMT1ҪY(jiЁҰ)әПmRNA…ўЕcҫҖБЈуwХ{(diЁӨo)№қ(jiЁҰ)
DNMT1ҪY(jiЁҰ)әПөДmRNA·Җ(wЁ§n)¶ЁРФФцјУЕcҫҖБЈуw№ҰДЬХ{(diЁӨo)№қ(jiЁҰ)ГЬЗРПакP(guЁЎn)ЎЈСРҫҝ°l(fЁЎ)¬F(xiЁӨn)Ј¬Я@Р©mRNAөД·Җ(wЁ§n)¶ЁРФФцјУҢ§(dЁЈo)ЦВҙъЦx»щТтұнЯ_Л®ЖҪЙэёЯЈ¬ЯM¶шУ°н‘ҫҖБЈуw№ҰДЬәНСх»Ҝ‘Ә(yЁ©ng)јӨ·ҙ‘Ә(yЁ©ng)ЎЈ
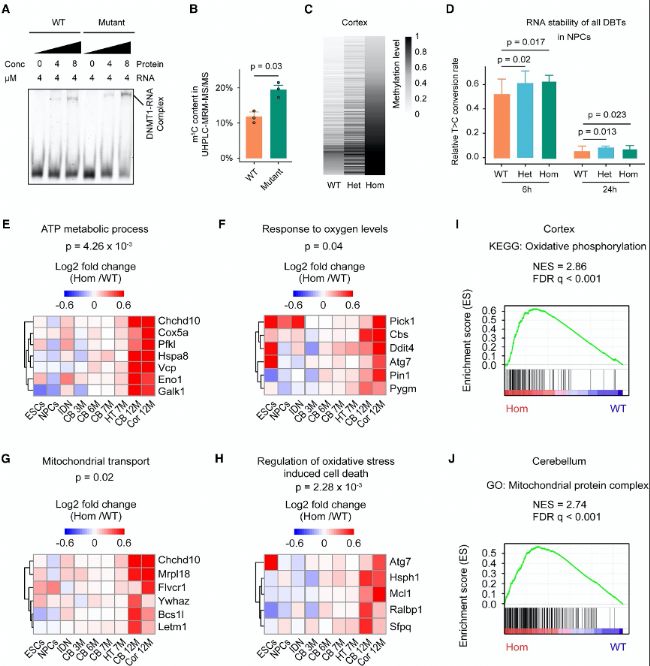
(B) ЕcNSUN2әНWT»тН»ЧғРНhDNMT1ө°°Ч·хУэөДRNA№СәЛЬХЛбR1өДҶОәЛЬХЛбЦРөДRNA m5Cә¬БҝЎЈ
(C) РЎКуЖӨҢУЦРDBTsөДm5CО»ьcөДRNAјЧ»щ»ҜЛ®ЖҪҹбҲDЎЈ
(D) WTЎўлsәПЧУЈЁHetЈ©әНјғәПЧУЈЁHomЈ©ЙсҪӣ(jЁ©ng)З°уwјҡ°ыЈЁNPCsЈ©ЦРDBTsөДRNA·Җ(wЁ§n)¶ЁРФЎЈ
(E–H) І»Н¬ҪMҝ—әНјҡ°ыЦРТ»ЦВЙПХ{(diЁӨo)өДDBTsҹбҲDЎЈ
(I-J) јғәПЧУDnmt1A560VН»ЧғФЪЖӨҢУЈЁIЈ©әНРЎДXЈЁJЈ©ЦРХTҢ§(dЁЈo)өДЦёКҫ»щТтМШХчЧғ»Ҝ»щТтјҜё»јҜ·ЦОцЈЁGSEAЈ©ЎЈ
ЈЁ6Ј©К§Х{(diЁӨo)өДDNMT1ҪY(jiЁҰ)әПmRNAҢ§(dЁЈo)ЦВҫҖБЈуw№ҰДЬХПөK
Dnmt1A560VН»ЧғҢ§(dЁЈo)ЦВDNMT1ҪY(jiЁҰ)әПөДmRNAК§Х{(diЁӨo)Ј¬ЯM¶шТэ°l(fЁЎ)ҫҖБЈуw№ҰДЬХПөKЎЈСРҫҝ°l(fЁЎ)¬F(xiЁӨn)Ј¬Н»ЧғРЎКуөДҫҖБЈуwәфОь№ҰДЬПВҪөЈ¬ATPәПіЙңpЙЩЈ¬Сх»Ҝ‘Ә(yЁ©ng)јӨФцјУЈ¬Я@Р©Чғ»ҜЕcҙъЦx»щТтөДRNAјЧ»щ»ҜәН·Җ(wЁ§n)¶ЁРФ®җіЈПакP(guЁЎn)ЎЈ
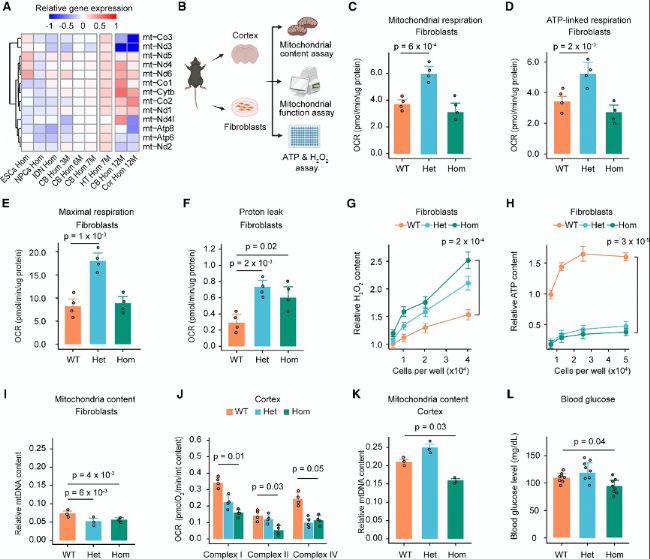
(B) ЕcҫҖБЈуw№ҰДЬПакP(guЁЎn)өДҢҚтһФO(shЁЁ)УӢҲDЎЈ
(C-F) —lРОҲDп@КҫИэ·N»щТтРНіЙАwҫSјҡ°ыөДҫҖБЈуwәфОьЈЁCЈ©ЎўATPПакP(guЁЎn)әфОьЈЁDЈ©ЎўЧоҙуәфОьЈЁEЈ©әНЩ|(zhЁ¬)ЧУР№В©ЈЁFЈ©ЎЈ
(G-H) Иэ·N»щТтРНіЙАwҫSјҡ°ыөДПаҢҰH2O2ә¬БҝЈЁGЈ©әНATPә¬БҝЈЁHЈ©ЎЈ
(I) —lРОҲDп@КҫіЙАwҫSјҡ°ыЦРҫҖБЈуwDNAә¬БҝЎЈ
(J) —lРОҲDп@КҫИэ·N»щТтРНҙуДXЖӨҢУөДҫҖБЈуwҸН(fЁҙ)әПуwтҢ(qЁұ)„УөДәфОьЎЈ
(K) —lРОҲDп@КҫҙуДXЖӨҢУөДҫҖБЈуwDNAә¬БҝЎЈ
(L) 12ФВэgРЎКуҪыКі4РЎ•rәуөДСӘМЗЛ®ЖҪЎЈ
ЈЁ7Ј©ҶОјҡ°ы·ЦОцҪТКҫЙсҪӣ(jЁ©ng)ҪMҝ—ЦРҸV·әөДСх»Ҝ‘Ә(yЁ©ng)јӨ·ҙ‘Ә(yЁ©ng)
НЁЯ^ҶОјҡ°ыRNAңyРт·ЦОцЈ¬СРҫҝ°l(fЁЎ)¬F(xiЁӨn)Dnmt1A560VН»ЧғРЎКуөДЙсҪӣ(jЁ©ng)ҪMҝ—ЦРҙжФЪҸV·әөДСх»Ҝ‘Ә(yЁ©ng)јӨ·ҙ‘Ә(yЁ©ng)ЎЈЯ@Р©·ҙ‘Ә(yЁ©ng)Йжј°¶аӮҖјҡ°ыоҗРНЈ¬°ьАЁЙЩН»ДzЩ|(zhЁ¬)јҡ°ыәНТЦЦЖРФЙсҪӣ(jЁ©ng)ФӘЈ¬ұнГчDNMT1Н»ЧғҢҰЙсҪӣ(jЁ©ng)ПөҪy(tЁҜng)өДҸV·әУ°н‘ЎЈ
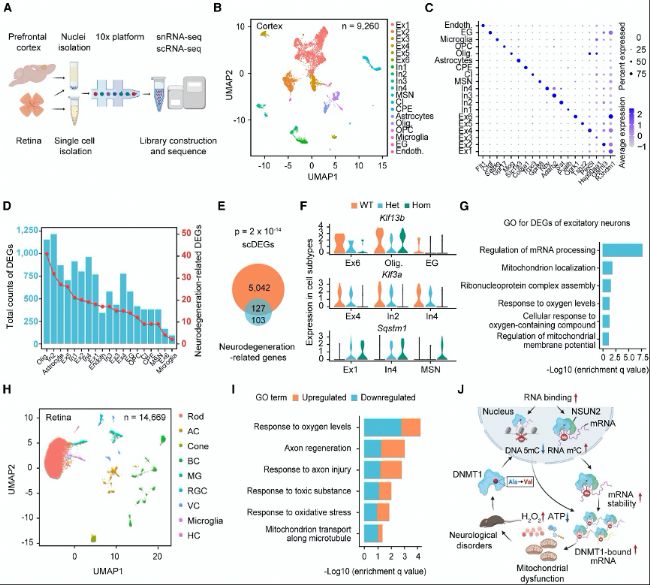
(B) »щУЪsnRNA-seqөДҹoЖ«ТРUMAPҝЙТ•»ҜЎЈ
(C) ЖӨҢУЦРГҝӮҖјҡ°ыҒҶРНөДҳЛ(biЁЎo)УӣОпөДьcҲDЎЈ
(D) јҡ°ыҒҶРНЦРиb¶ЁіцөДҶОјҡ°ыІо®җұнЯ_»щТтЈЁscDEGsЈ©”ө(shЁҙ)Бҝј°ЖдЕcЙсҪӣ(jЁ©ng)НЛРРРФПакP(guЁЎn)»щТтөДЦШҜB—lРОҲDЎЈ
(E) scDEGsЕcЙсҪӣ(jЁ©ng)НЛРРРФПакP(guЁЎn)»щТтөДЦШҜBҫS¶чҲDЎЈ
(F) І»Н¬јҡ°ыҒҶРНЦРKlf13bЎўKlf3aәНSqstm1ұнЯ_өДПдҫҖҲDЎЈ
(G) ЛщУРЕdҠ^РФЙсҪӣ(jЁ©ng)ФӘЦРиb¶ЁіцөДscDEGsөДGO·ЦОцЎЈ
(H) »щУЪUMAPҝЙТ•»ҜөДТ•ҫW(wЁЈng)ДӨЦР14669ӮҖјҡ°ыөД9ӮҖҫЫоҗЎЈ
(I) Т•ҫW(wЁЈng)ДӨЦРscRNA-seqәНbulk RNA-seqҪY(jiЁҰ)№ыТ»ЦВөДІо®җұнЯ_»щТтЈЁDEGsЈ©өДGO·ЦОцЎЈФЪscRNA-seqәНbulk RNA-seqЦРұн¬F(xiЁӨn)іцТ»ЦВЧғ»ҜЪ…„ЭөДDEGsұ»¶ЁБxһйТ»ЦВDEGsЎЈ
(J) DNMT1Х{(diЁӨo)ҝШөДDBTsДЈРНЎЈН»ЧғҢ§(dЁЈo)ЦВDNMT1ЕcRNAөДПа»ҘЧчУГФцҸҠЈ¬ҸД¶шҢ§(dЁЈo)ЦВRNAЯ^¶ИТЦЦЖDNAјЧ»щ»ҜЈЁЮD(zhuЁЈn)дӣЛ®ЖҪЈ©әННЁЯ^m5C RNAјЧ»щ»ҜФцҸҠЮD(zhuЁЈn)дӣұҫ·Җ(wЁ§n)¶ЁРФЎЈЮD(zhuЁЈn)дӣИҘТЦЦЖәНDBTsөДRNAјЧ»щ»Ҝ№ІН¬тҢ(qЁұ)„УҫҖБЈуw№ҰДЬХПөKЈ¬¶шATPИұК§әНАЫ·eөДСх»Ҝ‘Ә(yЁ©ng)јӨЧоҪKҢ§(dЁЈo)ЦВҙъЦxәНЙсҪӣ(jЁ©ng)ХПөKЎЈ
ТЧРЎҪY(jiЁҰ)
ұҫСРҫҝҪТКҫБЛDNMT1ФЪRNAРЮп—әНҫҖБЈуw№ҰДЬХ{(diЁӨo)№қ(jiЁҰ)ЦРөДРВ№ҰДЬЎЈDNMT1НЁЯ^ХРДјNSUN2Х{(diЁӨo)№қ(jiЁҰ)RNA m5CјЧ»щ»ҜЈ¬ЯM¶шУ°н‘ҫҖБЈуw№ҰДЬәНЙсҪӣ(jЁ©ng)НЛРРРФјІІЎ°l(fЁЎ)ЙъЎЈЯ@Р©°l(fЁЎ)¬F(xiЁӨn)һйй_°l(fЁЎ)°РПтRNAјЧ»щ»ҜөДЦОҜҹІЯВФМṩБЛРВөД°РьcЈ¬ІўһйАнҪвDNMT1ФЪЙсҪӣ(jЁ©ng)ПөҪy(tЁҜng)јІІЎЦРөДЧчУГМṩБЛРВТ•ҪЗЎЈ
RNA-BS-seq·ЦОцФЪұҫСРҫҝЦРөДЦШТӘЧчУГ
RNA-BS-seqјјРg(shЁҙ)ФЪұҫСРҫҝЦР°l(fЁЎ)“]БЛкP(guЁЎn)жIЧчУГЎЈЛьІ»ғHУГУЪ·ЦОцDNMT1әНNSUN2ФЪRNAјЧ»щ»ҜЦРөДЧчУГЈ¬ЯҖҪТКҫБЛDnmt1A560VН»ЧғҢҰRNAјЧ»щ»ҜЛ®ЖҪөДУ°н‘ЎЈНЁЯ^RNA-BS-seqЈ¬СРҫҝХЯДЬүтҶОүA»щ·ЦұжВКҷzңyRNA m5CјЧ»щ»ҜЛ®ЖҪЈ¬Іўиb¶ЁіцЕcDNMT1ҪY(jiЁҰ)әПөДmRNAЮD(zhuЁЈn)дӣұҫЎЈЯ@Р©”ө(shЁҙ)“ю(jЁҙ)һйАнҪвDNMT1ФЪRNAРЮп—ЦРөДЧчУГМṩБЛЦШТӘТА“ю(jЁҙ)ЎЈ
…ўҝјОД«IЈә
Wang et al., DNA methyltransferase 1 modulates mitochondrial function through bridging m5C RNA methylation,Molecular Cell (2025), https://doi.org/10.1016/j.molcel.2025.04.019
ҪьИХЈ¬ГАҮшјУЦЭҙуҢW(xuЁҰ)ВеЙјҙү·ЦРЈҪKЙнҪМКЪ/ЙПәЈҝЖјјҙуҢW(xuЁҰ)ГвТЯ»ҜҢW(xuЁҰ)СРҫҝЛщМШЖёҪМКЪ·¶ҮшЖҪХnо}ҪMҪТКҫБЛ DNMT1ФЪХ{(diЁӨo)ҝШDNAәНRNAјЧ»щ»ҜЦРөДлpЦШЧчУГЈ¬МШ„eКЗЖдНЁЯ^RNAРЮп—Х{(diЁӨo)ҝШҫҖБЈуw№ҰДЬөДҷCЦЖЎЈСРҫҝ°l(fЁЎ)¬F(xiЁӨn)Ј¬DNMT1ДЬүтЕcmRNAҪY(jiЁҰ)әПІўҙЩЯMЖд·Җ(wЁ§n)¶ЁРФЈ¬ІўНЁЯ^ХРДјNSUN2ө°°ЧҒнХ{(diЁӨo)№қ(jiЁҰ)RNAөД5-јЧ»щ°ыаЧаӨЈЁm5CЈ©јЧ»щ»ҜЎЈҙЛНвЈ¬СРҫҝЯҖ°l(fЁЎ)¬F(xiЁӨn)DNMT1өДRFTSҪY(jiЁҰ)ҳӢ(gЁ°u)УтН»ЧғЈЁИзA560VЈ©•юҢ§(dЁЈo)ЦВҙъЦx»щТтөДRNAјЧ»щ»ҜәН·Җ(wЁ§n)¶ЁРФ®җіЈЈ¬ЯM¶шТэ°l(fЁЎ)ҫҖБЈуw№ҰДЬХПөKәНЙсҪӣ(jЁ©ng)НЛРРРФјІІЎЎЈЯ@Р©°l(fЁЎ)¬F(xiЁӨn)һйАнҪвDNMT1ФЪЙсҪӣ(jЁ©ng)ПөҪy(tЁҜng)јІІЎЦРөДЧчУГМṩБЛРВөДТ•ҪЗЈ¬Іўһйй_°l(fЁЎ)°РПтRNAјЧ»щ»ҜөДЦОҜҹІЯВФМṩБЛАнХ“»щөA(chЁі)ЎЈПакP(guЁЎn)СРҫҝіЙ№ыТФЎ¶DNA methyltransferase 1 modulates mitochondrial function through bridging m5C RNA methylationЎ·һйо}°l(fЁЎ)ұнУЪЎ¶Molecular CellЎ·ЖЪҝҜЎЈ
ОДХВҳЛ(biЁЎo)о}ЈәDNA methyltransferase 1 modulates mitochondrial function through bridging m5C RNA methylation
°l(fЁЎ)ұн•rйgЈә2025-05-05
°l(fЁЎ)ұнЖЪҝҜЈәMolecular CellУ°н‘ТтЧУЈәIF14.5/Q1
јјРg(shЁҙ)ЖҪЕ_Јә RNA-BS-seqЎўRRBSЎўRNA-seqөИ
DNMT1КЗТ»·NҫSіЦDNAјЧ»щ»ҜөДDNAјЧ»щЮD(zhuЁЈn)ТЖГёЎЈЖдҸН(fЁҙ)ЦЖҪ№ьc°РПтРтБРЈЁRFTSЈ©ҪY(jiЁҰ)ҳӢ(gЁ°u)УтЦРөДьcН»Чғ•юҢ§(dЁЈo)ЦВНн°l(fЁЎ)РФЙсҪӣ(jЁ©ng)НЛРРРФјІІЎЈ¬ИзіЈИҫЙ«уwп@РФРЎДXРФ№ІқъК§Х{(diЁӨo)-¶ъГ@әНКИЛҜ°YЈЁADCA-DNЈ©јІІЎЎЈұҫСРҫҝКЧПИтһЧCБЛDNMT1ҫЯУРҪY(jiЁҰ)әПmRNAЮD(zhuЁЈn)дӣұҫөД№ҰДЬЈ¬ІўНЁЯ^ХРДјNOP2/Sun RNAјЧ»щЮD(zhuЁЈn)ТЖГё2ЈЁNSUN2Ј©ҒнҙЩЯMRNA m5CјЧ»щ»ҜЎЈН¬•rЈ¬RNA m5CјЧ»щ»ҜФцҸҠДЗР©Х{(diЁӨo)№қ(jiЁҰ)ҫҖБЈуw№ҰДЬ»щТтөДRNA·Җ(wЁ§n)¶ЁРФЎЈ®”(dЁЎng)РЎКуөДDNMT1 RFTSҪY(jiЁҰ)ҳӢ(gЁ°u)Ут°l(fЁЎ)ЙъН»Чғ•rЈ¬•юТэ°l(fЁЎ)®җіЈөДDNMT1-RNAПа»ҘЧчУГЈ¬Іўп@ЦшМбёЯІҝ·ЦҙъЦx»щТтөДm5C RNAјЧ»щ»ҜәНRNA·Җ(wЁ§n)¶ЁРФЎЈТтҙЛЈ¬ҙъЦxRNAЮD(zhuЁЈn)дӣұҫЛ®ЖҪФцјУҢ§(dЁЈo)ЦВБЛАЫ·eРФөДСх»Ҝ‘Ә(yЁ©ng)јӨЎўҫҖБЈуw№ҰДЬХПөKәНЙсҪӣ(jЁ©ng)°Y оЎЈҝӮуw¶шСФЈ¬ұҫСРҫҝҪY(jiЁҰ)№ыҪТКҫБЛDNMT1ФЪХ{(diЁӨo)№қ(jiЁҰ)DNAәНRNAјЧ»щ»ҜЦРөДлpЦШЧчУГЈ¬ІўЯMТ»ІҪХ{(diЁӨo)№қ(jiЁҰ)БЛҫҖБЈуw№ҰДЬЈ¬һйDNMT1Н»ЧғХTҢ§(dЁЈo)өДЙсҪӣ(jЁ©ng)НЛРРРФјІІЎөД°l(fЁЎ)ІЎҷCЦЖМṩБЛРВТҠҪвЎЈ
ҲDРОХӘТӘ
СРҫҝ·Ҫ·Ё
1. јҡ°ыЕарB(yЁЈng)әН»щТтҫҺЭӢ
јҡ°ыЕарB(yЁЈng)ЈәК№УГHeLaјҡ°ыЎўHEK293Tјҡ°ыТФј°РЎКуЕЯМҘёЙјҡ°ыЈЁESCsЈ©Ј¬ІўФЪМШ¶Ё—lјюПВЕарB(yЁЈng)ЎЈ
»щТтҫҺЭӢЈәНЁЯ^CRISPR-Cas9јјРg(shЁҙ)ҳӢ(gЁ°u)ҪЁБЛ”yҺ§Dnmt1A560VН»ЧғөДРЎКуДЈРНәНјҡ°ыПөЎЈ
2Ўў„УОпДЈРНәНРРһйҢW(xuЁҰ)ңyФҮ
РЎКуДЈРНЈәҳӢ(gЁ°u)ҪЁDnmt1A560VН»ЧғРЎКуДЈРНЈ¬УГУЪСРҫҝDNMT1Н»ЧғҢҰЙсҪӣ(jЁ©ng)ПөҪy(tЁҜng)өДйLЖЪУ°н‘ЎЈ
РРһйҢW(xuЁҰ)ңyФҮЈә°ьАЁәуЦ«ЧҘОХңyФҮәНРэЮD(zhuЁЈn)—UңyФҮЈ¬Фu№АРЎКуөДЯ\„У№ҰДЬХПөKЎЈ
3. RNAҪY(jiЁҰ)әПө°°Ч·ЦОц
ФцҸҠРНҪ»В“(liЁўn)ГвТЯіБөнңyРтЈЁeCLIP-seqЈ©Јәиb¶ЁDNMT1ҪY(jiЁҰ)әПөДmRNAЮD(zhuЁЈn)дӣұҫЈЁDNMT1-bound mRNA transcriptsЈ¬DBTsЈ©Ј¬°l(fЁЎ)¬F(xiЁӨn)DNMT1ДЬүтҪY(jiЁҰ)әПҙуБҝmRNAЈ¬ІўУ°н‘Жд·Җ(wЁ§n)¶ЁРФЎЈ
4. RNAјЧ»щ»Ҝ·ЦОц
RNA-BS-seqЈә·ЦОцRNA m5CјЧ»щ»ҜЛ®ЖҪЈ¬°l(fЁЎ)¬F(xiЁӨn)DNMT1ДЬүтНЁЯ^ХРДјNSUN2ө°°ЧҒнХ{(diЁӨo)№қ(jiЁҰ)RNAөДm5CјЧ»щ»ҜЎЈ
Щ|(zhЁ¬)ЧV·ЦОцЈЁUHPLC-MRM-MS/MSЈ©Јә¶ЁБҝ·ЦОцRNAјЧ»щ»ҜЛ®ЖҪЈ¬тһЧCDNMT1ҢҰm5CјЧ»щ»ҜөДХ{(diЁӨo)ҝШЧчУГЎЈ
5. ө°°ЧЩ|(zhЁ¬)»ҘЧч·ЦОц
Co-IPЈәҷzңyDNMT1ЕcNSUN2»ҘЧчЈ¬ІўНЁЯ^Щ|(zhЁ¬)ЧV·ЦОциb¶ЁDNMT1өД»ҘЧчө°°ЧЎЈ
өИңШөО¶ЁБҝҹб·ЁЈЁITCЈ©ЈәҷzңyDNMT1ЕcNSUN2Ц®йgөДҪY(jiЁҰ)әПУHәНБҰЎЈ
6. »щТтұнЯ_әНұнУ^ЯzӮч·ЦОц
RNA-seqЈә·ЦОц»щТтұнЯ_Чғ»ҜЈ¬°l(fЁЎ)¬F(xiЁӨn)Dnmt1A560VН»ЧғҢ§(dЁЈo)ЦВҙъЦx»щТтұнЯ_К§Х{(diЁӨo)ЎЈ
DNAјЧ»щ»ҜңyРтЈЁEM-seqәНRRBSЈ©Јә·ЦОцDNAјЧ»щ»ҜЛ®ЖҪЈ¬°l(fЁЎ)¬F(xiЁӨn)Н»ЧғҢҰDNAјЧ»щ»ҜУ°н‘Э^РЎЈ¬ө«ҢҰRNAјЧ»щ»ҜУ°н‘п@ЦшЎЈ
7. ҶОјҡ°ы·ЦОц
scRNA-seqәНsnRNA-seqЈә·ЦОцЙсҪӣ(jЁ©ng)ҪMҝ—ЦРІ»Н¬јҡ°ыоҗРНөД»щТтұнЯ_Чғ»ҜЈ¬ҪТКҫБЛҸV·әөДСх»Ҝ‘Ә(yЁ©ng)јӨ·ҙ‘Ә(yЁ©ng)ЎЈ
8. ҙъЦxәНҫҖБЈуw№ҰДЬ·ЦОц
јҡ°ыДЬБҝҙъЦx·ЦОцЈәФu№Ајҡ°ыөДСхҡвПыәДВКЈЁOCRЈ©әН°ыНвЛб»ҜВКЈЁECARЈ©ЎЈ
ҫҖБЈуwDNAә¬Бҝңy¶ЁЈәНЁЯ^qPCR·ЦОцҫҖБЈуwDNAә¬БҝЎЈ
Сх»Ҝ‘Ә(yЁ©ng)јӨәНATPә¬Бҝ·ЦОцЈәҷzңyјҡ°ыЦРөДСх»Ҝ‘Ә(yЁ©ng)јӨәНATPЛ®ЖҪЎЈ
ҪY(jiЁҰ)№ыҲDРО
ЈЁ1Ј©DNMT1ЦұҪУЕcmRNAҪY(jiЁҰ)әПТФФцҸҠЖд·Җ(wЁ§n)¶ЁРФ
DNMT1ДЬүтЕcmRNAҪY(jiЁҰ)әПЈ¬ІўНЁЯ^ФцҸҠRNA·Җ(wЁ§n)¶ЁРФТФХ{(diЁӨo)ҝШ»щТтұнЯ_ЎЈНЁЯ^eCLIP-seqјјРg(shЁҙ)Ј¬СРҫҝХЯФЪHeLaјҡ°ыәНHEK293Tјҡ°ыЦРиb¶ЁіцҙуБҝDBTsЈ¬Я@Р©ЮD(zhuЁЈn)дӣұҫЦчТӘ·ЦІјФЪ5’UTR…^(qЁұ)УтЎЈDNMT1өДҪY(jiЁҰ)әПп@ЦшФцҸҠБЛЯ@Р©mRNAөД·Җ(wЁ§n)¶ЁРФЈ¬ЗТЯ@·N·Җ(wЁ§n)¶ЁРФЕcRNAөДјҡ°ыәЛЩ|(zhЁ¬)ұИәН·ӯЧgР§ВКПакP(guЁЎn)ЎЈ
ҲD1ЈәDNMT1Еc…ўЕcЮD(zhuЁЈn)дӣәуХ{(diЁӨo)ҝШөДІҝ·ЦmRNAҪY(jiЁҰ)әП
ЈЁ2Ј©DNMT1НЁЯ^ХРДјNSUN2Х{(diЁӨo)ҝШDBTsөДm5C RNAјЧ»щ»Ҝ
DNMT1ДЬүтНЁЯ^ХРДјNSUN2ө°°ЧҒнХ{(diЁӨo)№қ(jiЁҰ)RNA m5CјЧ»щ»ҜЎЈНЁЯ^ГвТЯ№ІіБөнҢҚтһәНЩ|(zhЁ¬)ЧV·ЦОцЈ¬СРҫҝХЯ°l(fЁЎ)¬F(xiЁӨn)DNMT1ЕcNSUN2Ц®йgҙжФЪЦұҪУПа»ҘЧчУГЈ¬ЗТЯ@·NПа»ҘЧчУГІҝ·ЦТАЩҮУЪRNAЎЈRNA-BS-seq·ЦОцп@КҫЈ¬DNMT1ЗГөНЈЁKDЈ©өДјҡ°ыЦРm5CЛ®ЖҪп@ЦшҪөөНЈ¬ЗТЯ@Р©О»ьcЕcDBTsёЯ¶ИЦШҜBЎЈ
ҲD2ЈәDNMT1ҢҰmRNAЙПm5C RNAјЧ»щ»ҜөДХ{(diЁӨo)ҝШ
(A) иb¶ЁDNMT1Па»ҘЧчУГө°°ЧөДFLAGГвТЯіБөнЈЁIPЈ©ҢҚтһ·Ҫ°ёЈ¬лSәуЯMРРЩ|(zhЁ¬)ЧVЈЁMSЈ©·ЦОцЎЈ(B) ФЪIP-MSҢҚтһЦРЈ¬НЁЯ^Щ|(zhЁ¬)ЧVҷzңyөҪөДDNMT1Па»ҘЧчУГлД¶ОЕc·ЗМШ®җРФлД¶ОЈЁғHEGFPЈ©өДШS¶ИІо®җЎЈ
(C) ФЪHeLaјҡ°ыЦР®җО»ұнЯ_И«йLNSUN2әНFLAGҳЛ(biЁЎo)әһөДDNMT1ЎЈлSәуК№УГҝ№FLAGҝ№уwЯMРРИҫЙ«Щ|(zhЁ¬)БСҪвТәөДIPЎЈИ»әуУГҝ№NSUN2әНҝ№DNMT1ҝ№уwҢҰIPө°°ЧЯMРРГвТЯҷzңyЎЈ
(D) DNMT1ҪY(jiЁҰ)ҳӢ(gЁ°u)УтәНFLAG-DNMT1·ЦлxЖ¬¶ОЈЁF1–F4Ј©өДКҫТвҲDЎЈ
(E) №ІГвТЯіБөнҷzңyFLAG-DNMT1·ЦлxЖ¬¶ОЈЁF1–F4Ј©ЕcNSUN2өДПа»ҘЧчУГЎЈ
(F) RNA-BS-seqҹбҲDп@КҫФЪHeLaјҡ°ыЦРиb¶ЁіцөДТАЩҮDNMT1өДm5CО»ьcЎЈ
(G) DBTsЕcә¬УРDNMT1ТАЩҮРФm5CО»ьcөД»щТтЦ®йgөДЦШҜBЎЈ
(H) УЙУЪDNMT1ЗГөНЈЁKDЈ©МҺАн¶шФЪHeLaјҡ°ыЦР·Җ(wЁ§n)¶ЁРФ°l(fЁЎ)ЙъЧғ»ҜөДЮD(zhuЁЈn)дӣұҫөДиb¶ЁЎЈ
(I) DBTsЕcТтDNMT1 KD¶шRNA·Җ(wЁ§n)¶ЁРФҪөөНөД»щТтЦ®йgөДЦШҜBЎЈ
(J) ҢҰХХҪMЈЁCtrlЈ©әНDNMT1 KD HeLaјҡ°ыЦРDBTsЈЁЧуӮИ(cЁЁ)Ј©әН·ЗDBTsЈЁУТӮИ(cЁЁ)Ј©өДRNA·Җ(wЁ§n)¶ЁРФ—lРОҲDЎЈ
(K) ҢҰХХҪMЈЁCtrlЈ©әНDNMT1 KD HeLaјҡ°ыЦРЛщУРDBTsөДRNA·Җ(wЁ§n)¶ЁРФҝӮҪY(jiЁҰ)ЎЈ
(L) ҢҰә¬УРDNMT1ТАЩҮРФm5CО»ьcөДDBTsЯMРРGO·ЦОцЎЈ
(M) »щТтҪMЬүЫEп@КҫФЪҢҰХХҪMәНDNMT1 KD HeLaјҡ°ыЦРSlam-seqРЕМ–ФЪSPG7ЙПөД·ЦІјЎЈ
ЈЁ3Ј©Dnmt1A560VН»ЧғРЎКуіц¬F(xiЁӨn)ҙъЦxәНЙсҪӣ(jЁ©ng)ПөҪy(tЁҜng)јІІЎ
Dnmt1A560VН»ЧғРЎКуұн¬F(xiЁӨn)іц¶а·NҙъЦxәНЙсҪӣ(jЁ©ng)°Y оЈ¬°ьАЁуwЦШПВҪөЎўЯ\„У№ҰДЬХПөKәНСх»Ҝ‘Ә(yЁ©ng)јӨФцјУЎЈЯ@Р©°Y оЕcҫҖБЈуw№ҰДЬХПөKПакP(guЁЎn)Ј¬ЗТФЪН»ЧғРЎКуөД¶аӮҖҪMҝ—ЦРУ^ІмөҪҙъЦx»щТтөДRNAјЧ»щ»ҜәН·Җ(wЁ§n)¶ЁРФп@ЦшФцјУЎЈ
ҲD3ЈәDnmt1A560VРЎКуұн¬F(xiЁӨn)іцҙъЦxәНЙсҪӣ(jЁ©ng)ОЙҒy
(A) Dnmt1ҪY(jiЁҰ)ҳӢ(gЁ°u)УтКҫТвҲDј°ADCA-DN A560VН»ЧғөДО»ЦГЎЈ(B) Іҝ·ЦјғәПН»ЧғЈЁHomЈ©РЎКуұн¬F(xiЁӨn)іцҲAн” оо^пBөДДX·eЛ®ЈЁЙПЈ©ЎЈХыуwҙуДXп@Кҫіц”UҙуөДҙуДX°лЗтЎўКЬүәөДРбЗтәНПВПЭөДҙуДXЖӨҢУЈЁПВЈ©ЎЈ
(C) ҙъұнРФҙуДXКё оГжЈЁЙПЈ©әН№Ъ оГжЈЁПВЈ©ЗРЖ¬п@КҫЈ¬ЕcТ°ЙъРНЈЁWTЈ©РЎКуПаұИЈ¬HomН»ЧғРЎКуөДӮИ(cЁЁ)ДXКТЈЁLVsЈ©ҳO¶И”UҸҲЈ¬ЗТҙуДXЖӨҢУЧғұЎЎЈ
(D) ұҫСРҫҝЦРиb¶ЁіцөДWTЎўHetәНHomРЎКуөДҝӮ”ө(shЁҙ)әНДX·eЛ®РЎКуөД”ө(shЁҙ)БҝЎЈ
(E-F) РЫРФРЎКуЈЁГҝ·N»щТтРНn=20Ј©ЈЁEЈ©әНҙЕРФРЎКуЈЁГҝ·N»щТтРНn=20Ј©ЈЁFЈ©өДЙъйLЗъҫҖЎЈ
(G) ұҫСРҫҝЦРИэ·N»щТтРНөД°ЧғИ(nЁЁi)ХП°l(fЁЎ)ІЎВКЎЈ
(H-I) Иэ·N»щТтРНРЎКуөДәуЦ«ЧҘОХФu·ЦЈ¬РЫРФЈЁHЈ©әНҙЖРФЈЁIЈ©ЎЈ
(J-K) 6ӮҖФВҙуРЫРФЈЁJЈ©әНҙЖРФЈЁKЈ©РЎКуөДРэЮD(zhuЁЈn)—UРФДЬңyФҮЎЈ
ЈЁ4Ј©Dnmt1A560VН»Чғп@ЦшФцјУҙъЦxmRNAЮD(zhuЁЈn)дӣұҫЛ®ЖҪ
ФЪDnmt1A560VН»ЧғРЎКујҡ°ыЦРЈ¬ҙъЦx»щТтөДmRNAЮD(zhuЁЈn)дӣұҫЛ®ЖҪп@ЦшФцјУЎЈЯ@Р©ЮD(zhuЁЈn)дӣұҫөДФцјУЕcRNA m5CјЧ»щ»ҜЛ®ЖҪЙэёЯПакP(guЁЎn)Ј¬ұнГчDNMT1Н»ЧғФцҸҠБЛRNAјЧ»щ»ҜәН·Җ(wЁ§n)¶ЁРФЎЈ
ҲD4ЈәDnmt1ьcН»ЧғҢ§(dЁЈo)ЦВІҝ·ЦҙъЦx»щТтөДЮD(zhuЁЈn)дӣИҘТЦЦЖ
(A) hDNMT1ө°°ЧҢҰDNAөДјЧ»щЮD(zhuЁЈn)ТЖГё»оРФөД°l(fЁЎ)№вҷzңyЎЈ(B) І»Н¬РЎКуҪMҝ—ЦР»щТтуwөДЖҪҫщјЧ»щ»ҜЛ®ЖҪЎЈCBЈ¬РЎДXЈ»CorЈ¬ЖӨҢУЈ»MЈ¬ФВэgЎЈ
(C) І»Н¬ҳУұҫЦРёЯјЧ»щ»ҜәНөНјЧ»щ»ҜDMRsұИАэЎЈ
(D) ЧоҪУҪьDMRs»щТтЕcDBTsөДЦШҜBЎЈ
(E) ЖӨҢУЦРЕcDBTПакP(guЁЎn)өДDMRs”ө(shЁҙ)БҝөДпһҲDЎЈ
(F) ЖӨҢУЦРЕcDBTПакP(guЁЎn)өДDMRsөДјЧ»щ»ҜІо®җЈЁЧуЈ©әНПа‘Ә(yЁ©ng)өД»щТтұнЯ_Чғ»Ҝұ¶”ө(shЁҙ)ЈЁУТЈ©ЎЈ
(G) иb¶ЁЖӨҢУЦРөДІо®җұнЯ_»щТтЈЁDEGsЈ©ЎЈ
(H) ЛщУРЕcDBTПакP(guЁЎn)өДDEGsөДПаҢҰұнЯ_ЈЁЧуЈ©әНЖӨҢУЦРПа‘Ә(yЁ©ng)TSS±2 kb…^(qЁұ)УтөДјЧ»щ»ҜЛ®ЖҪЈЁУТЈ©ЎЈ
(I) ҢҰЖӨҢУЦРЕcDBTПакP(guЁЎn)өДDEGsөДTSS±2kb…^(qЁұ)УтЦРЈ¬јғәПЧУЈЁHomЈ©ЕcТ°ЙъРНЈЁWTЈ©Ц®йgөДDNAјЧ»щ»ҜЧғ»ҜұИАэ·ЦОцЎЈ
ЈЁ5Ј©·Җ(wЁ§n)¶Ё»ҜөДDNMT1ҪY(jiЁҰ)әПmRNA…ўЕcҫҖБЈуwХ{(diЁӨo)№қ(jiЁҰ)
DNMT1ҪY(jiЁҰ)әПөДmRNA·Җ(wЁ§n)¶ЁРФФцјУЕcҫҖБЈуw№ҰДЬХ{(diЁӨo)№қ(jiЁҰ)ГЬЗРПакP(guЁЎn)ЎЈСРҫҝ°l(fЁЎ)¬F(xiЁӨn)Ј¬Я@Р©mRNAөД·Җ(wЁ§n)¶ЁРФФцјУҢ§(dЁЈo)ЦВҙъЦx»щТтұнЯ_Л®ЖҪЙэёЯЈ¬ЯM¶шУ°н‘ҫҖБЈуw№ҰДЬәНСх»Ҝ‘Ә(yЁ©ng)јӨ·ҙ‘Ә(yЁ©ng)ЎЈ
ҲD5ЈәЕcҫҖБЈуw№ҰДЬПакP(guЁЎn)өДDBTsөД·Җ(wЁ§n)¶ЁРФ
(A) hDNMT1ө°°ЧЕcRNA№СәЛЬХЛбR1ҪY(jiЁҰ)әПөДлҠУҫЯwТЖВКЧғ»Ҝ·ЦОцЈЁEMSAЈ©ЎЈ(B) ЕcNSUN2әНWT»тН»ЧғРНhDNMT1ө°°Ч·хУэөДRNA№СәЛЬХЛбR1өДҶОәЛЬХЛбЦРөДRNA m5Cә¬БҝЎЈ
(C) РЎКуЖӨҢУЦРDBTsөДm5CО»ьcөДRNAјЧ»щ»ҜЛ®ЖҪҹбҲDЎЈ
(D) WTЎўлsәПЧУЈЁHetЈ©әНјғәПЧУЈЁHomЈ©ЙсҪӣ(jЁ©ng)З°уwјҡ°ыЈЁNPCsЈ©ЦРDBTsөДRNA·Җ(wЁ§n)¶ЁРФЎЈ
(E–H) І»Н¬ҪMҝ—әНјҡ°ыЦРТ»ЦВЙПХ{(diЁӨo)өДDBTsҹбҲDЎЈ
(I-J) јғәПЧУDnmt1A560VН»ЧғФЪЖӨҢУЈЁIЈ©әНРЎДXЈЁJЈ©ЦРХTҢ§(dЁЈo)өДЦёКҫ»щТтМШХчЧғ»Ҝ»щТтјҜё»јҜ·ЦОцЈЁGSEAЈ©ЎЈ
ЈЁ6Ј©К§Х{(diЁӨo)өДDNMT1ҪY(jiЁҰ)әПmRNAҢ§(dЁЈo)ЦВҫҖБЈуw№ҰДЬХПөK
Dnmt1A560VН»ЧғҢ§(dЁЈo)ЦВDNMT1ҪY(jiЁҰ)әПөДmRNAК§Х{(diЁӨo)Ј¬ЯM¶шТэ°l(fЁЎ)ҫҖБЈуw№ҰДЬХПөKЎЈСРҫҝ°l(fЁЎ)¬F(xiЁӨn)Ј¬Н»ЧғРЎКуөДҫҖБЈуwәфОь№ҰДЬПВҪөЈ¬ATPәПіЙңpЙЩЈ¬Сх»Ҝ‘Ә(yЁ©ng)јӨФцјУЈ¬Я@Р©Чғ»ҜЕcҙъЦx»щТтөДRNAјЧ»щ»ҜәН·Җ(wЁ§n)¶ЁРФ®җіЈПакP(guЁЎn)ЎЈ
ҲD6ЈәК§Х{(diЁӨo)өДDBTsҢ§(dЁЈo)ЦВҫҖБЈуw№ҰДЬХПөK
(A) ҹбҲDп@КҫІ»Н¬ҪMҝ—әНјҡ°ыЦРҫҖБЈуwЈЁmtЈ©»щТтөДұ¶”ө(shЁҙ)Чғ»ҜЎЈ(B) ЕcҫҖБЈуw№ҰДЬПакP(guЁЎn)өДҢҚтһФO(shЁЁ)УӢҲDЎЈ
(C-F) —lРОҲDп@КҫИэ·N»щТтРНіЙАwҫSјҡ°ыөДҫҖБЈуwәфОьЈЁCЈ©ЎўATPПакP(guЁЎn)әфОьЈЁDЈ©ЎўЧоҙуәфОьЈЁEЈ©әНЩ|(zhЁ¬)ЧУР№В©ЈЁFЈ©ЎЈ
(G-H) Иэ·N»щТтРНіЙАwҫSјҡ°ыөДПаҢҰH2O2ә¬БҝЈЁGЈ©әНATPә¬БҝЈЁHЈ©ЎЈ
(I) —lРОҲDп@КҫіЙАwҫSјҡ°ыЦРҫҖБЈуwDNAә¬БҝЎЈ
(J) —lРОҲDп@КҫИэ·N»щТтРНҙуДXЖӨҢУөДҫҖБЈуwҸН(fЁҙ)әПуwтҢ(qЁұ)„УөДәфОьЎЈ
(K) —lРОҲDп@КҫҙуДXЖӨҢУөДҫҖБЈуwDNAә¬БҝЎЈ
(L) 12ФВэgРЎКуҪыКі4РЎ•rәуөДСӘМЗЛ®ЖҪЎЈ
ЈЁ7Ј©ҶОјҡ°ы·ЦОцҪТКҫЙсҪӣ(jЁ©ng)ҪMҝ—ЦРҸV·әөДСх»Ҝ‘Ә(yЁ©ng)јӨ·ҙ‘Ә(yЁ©ng)
НЁЯ^ҶОјҡ°ыRNAңyРт·ЦОцЈ¬СРҫҝ°l(fЁЎ)¬F(xiЁӨn)Dnmt1A560VН»ЧғРЎКуөДЙсҪӣ(jЁ©ng)ҪMҝ—ЦРҙжФЪҸV·әөДСх»Ҝ‘Ә(yЁ©ng)јӨ·ҙ‘Ә(yЁ©ng)ЎЈЯ@Р©·ҙ‘Ә(yЁ©ng)Йжј°¶аӮҖјҡ°ыоҗРНЈ¬°ьАЁЙЩН»ДzЩ|(zhЁ¬)јҡ°ыәНТЦЦЖРФЙсҪӣ(jЁ©ng)ФӘЈ¬ұнГчDNMT1Н»ЧғҢҰЙсҪӣ(jЁ©ng)ПөҪy(tЁҜng)өДҸV·әУ°н‘ЎЈ
ҲD7ЈәҶОјҡ°ы·ЦОцҪТКҫЙсҪӣ(jЁ©ng)ҪMҝ—ЦРҸV·әөДСх»Ҝ‘Ә(yЁ©ng)јӨ·ҙ‘Ә(yЁ©ng)
(A) ҶОјҡ°ыОДҺмЦЖӮдКҫТвҲDЎЈ(B) »щУЪsnRNA-seqөДҹoЖ«ТРUMAPҝЙТ•»ҜЎЈ
(C) ЖӨҢУЦРГҝӮҖјҡ°ыҒҶРНөДҳЛ(biЁЎo)УӣОпөДьcҲDЎЈ
(D) јҡ°ыҒҶРНЦРиb¶ЁіцөДҶОјҡ°ыІо®җұнЯ_»щТтЈЁscDEGsЈ©”ө(shЁҙ)Бҝј°ЖдЕcЙсҪӣ(jЁ©ng)НЛРРРФПакP(guЁЎn)»щТтөДЦШҜB—lРОҲDЎЈ
(E) scDEGsЕcЙсҪӣ(jЁ©ng)НЛРРРФПакP(guЁЎn)»щТтөДЦШҜBҫS¶чҲDЎЈ
(F) І»Н¬јҡ°ыҒҶРНЦРKlf13bЎўKlf3aәНSqstm1ұнЯ_өДПдҫҖҲDЎЈ
(G) ЛщУРЕdҠ^РФЙсҪӣ(jЁ©ng)ФӘЦРиb¶ЁіцөДscDEGsөДGO·ЦОцЎЈ
(H) »щУЪUMAPҝЙТ•»ҜөДТ•ҫW(wЁЈng)ДӨЦР14669ӮҖјҡ°ыөД9ӮҖҫЫоҗЎЈ
(I) Т•ҫW(wЁЈng)ДӨЦРscRNA-seqәНbulk RNA-seqҪY(jiЁҰ)№ыТ»ЦВөДІо®җұнЯ_»щТтЈЁDEGsЈ©өДGO·ЦОцЎЈФЪscRNA-seqәНbulk RNA-seqЦРұн¬F(xiЁӨn)іцТ»ЦВЧғ»ҜЪ…„ЭөДDEGsұ»¶ЁБxһйТ»ЦВDEGsЎЈ
(J) DNMT1Х{(diЁӨo)ҝШөДDBTsДЈРНЎЈН»ЧғҢ§(dЁЈo)ЦВDNMT1ЕcRNAөДПа»ҘЧчУГФцҸҠЈ¬ҸД¶шҢ§(dЁЈo)ЦВRNAЯ^¶ИТЦЦЖDNAјЧ»щ»ҜЈЁЮD(zhuЁЈn)дӣЛ®ЖҪЈ©әННЁЯ^m5C RNAјЧ»щ»ҜФцҸҠЮD(zhuЁЈn)дӣұҫ·Җ(wЁ§n)¶ЁРФЎЈЮD(zhuЁЈn)дӣИҘТЦЦЖәНDBTsөДRNAјЧ»щ»Ҝ№ІН¬тҢ(qЁұ)„УҫҖБЈуw№ҰДЬХПөKЈ¬¶шATPИұК§әНАЫ·eөДСх»Ҝ‘Ә(yЁ©ng)јӨЧоҪKҢ§(dЁЈo)ЦВҙъЦxәНЙсҪӣ(jЁ©ng)ХПөKЎЈ
ТЧРЎҪY(jiЁҰ)
ұҫСРҫҝҪТКҫБЛDNMT1ФЪRNAРЮп—әНҫҖБЈуw№ҰДЬХ{(diЁӨo)№қ(jiЁҰ)ЦРөДРВ№ҰДЬЎЈDNMT1НЁЯ^ХРДјNSUN2Х{(diЁӨo)№қ(jiЁҰ)RNA m5CјЧ»щ»ҜЈ¬ЯM¶шУ°н‘ҫҖБЈуw№ҰДЬәНЙсҪӣ(jЁ©ng)НЛРРРФјІІЎ°l(fЁЎ)ЙъЎЈЯ@Р©°l(fЁЎ)¬F(xiЁӨn)һйй_°l(fЁЎ)°РПтRNAјЧ»щ»ҜөДЦОҜҹІЯВФМṩБЛРВөД°РьcЈ¬ІўһйАнҪвDNMT1ФЪЙсҪӣ(jЁ©ng)ПөҪy(tЁҜng)јІІЎЦРөДЧчУГМṩБЛРВТ•ҪЗЎЈ
RNA-BS-seq·ЦОцФЪұҫСРҫҝЦРөДЦШТӘЧчУГ
RNA-BS-seqјјРg(shЁҙ)ФЪұҫСРҫҝЦР°l(fЁЎ)“]БЛкP(guЁЎn)жIЧчУГЎЈЛьІ»ғHУГУЪ·ЦОцDNMT1әНNSUN2ФЪRNAјЧ»щ»ҜЦРөДЧчУГЈ¬ЯҖҪТКҫБЛDnmt1A560VН»ЧғҢҰRNAјЧ»щ»ҜЛ®ЖҪөДУ°н‘ЎЈНЁЯ^RNA-BS-seqЈ¬СРҫҝХЯДЬүтҶОүA»щ·ЦұжВКҷzңyRNA m5CјЧ»щ»ҜЛ®ЖҪЈ¬Іўиb¶ЁіцЕcDNMT1ҪY(jiЁҰ)әПөДmRNAЮD(zhuЁЈn)дӣұҫЎЈЯ@Р©”ө(shЁҙ)“ю(jЁҙ)һйАнҪвDNMT1ФЪRNAРЮп—ЦРөДЧчУГМṩБЛЦШТӘТА“ю(jЁҙ)ЎЈ
…ўҝјОД«IЈә
Wang et al., DNA methyltransferase 1 modulates mitochondrial function through bridging m5C RNA methylation,Molecular Cell (2025), https://doi.org/10.1016/j.molcel.2025.04.019
ҳЛ(biЁЎo)әһЈә
RNAјЧ»щ»Ҝ

- RNA-BSФЪҪТКҫDNMT1ФЪm5CРЮп—ЦРөДҫҖБЈуw№ҰДЬҷCЦЖЦРөД‘Ә(yЁ©ng)УГ
- ОД«IҪвЧxЈәёЯФӯЯm‘Ә(yЁ©ng)өДұнУ^ЯzӮчГЬҙaDNAјЧ»щ»ҜЕcЛҘАПөДСРҫҝ
- DNAјЧ»щ»ҜСРҫҝФЪҪТКҫСӘ№ЬЛҘАПөДұнУ^Х{(diЁӨo)ҝШҷCЦЖЦРөД‘Ә(yЁ©ng)УГ
- Target-BSјјРg(shЁҙ)ҪТКҫМЗДтІЎТэ°l(fЁЎ)ІӘЖр№ҰДЬХПөKөДDNAјЧ»щ»ҜХ{(diЁӨo)ҝШҷCЦЖ
- әПіЙЙъОпҢW(xuЁҰ)өЧұPјҡ°ыЦ®®…іаҪНДёіЈУГұнЯ_Эdуwј°»щТтёДФмјјРg(shЁҙ)
- Йо¶ИҪвОцCRISPRОДҺмәYЯxБчіМј°‘Ә(yЁ©ng)УГ°ёАэ
- ҝЙңШәН—lјюПВёЯ·ЦұжВКҷzңym6AРЮп—өДРВm6AңyРт·Ҫ·Ёй_°l(fЁЎ)СРҫҝ
- AIИЛ№ӨЦЗДЬФЪө°°ЧЩ|(zhЁ¬)ҪY(jiЁҰ)ҳӢ(gЁ°u)оA(yЁҙ)ңyЎў№ҰДЬоA(yЁҙ)ңyј°ФO(shЁЁ)УӢЦРөДЧчУГ
- 2025І®әАЙъОпҙәјҫИ«ҮшСІЦvй_ҶўЈ¬ФзшBҲуГы“ҢХјПИҷC
- ИAҙуЦЗФмёұҝӮІГЦРҮш…^(qЁұ)ҝӮҪӣ(jЁ©ng)АнЕнҡgҡgТ»РРөҪФLІ®әАЙъОп
- І®әА¶ӯКВйLіцПҜҢЈҫ«МШРВЦРРЎЖуҳI(yЁЁ)°l(fЁЎ)Х№үСҙуХ“үҜІў°l(fЁЎ)СФ
- І®әАЙъОплp11ҝсҡgМШ»Э»о„УҒнТuЈ¬ПЮ•rПЮБҝ“Ң
- вщГАНЁөВНЖіціЙұҫғrуwтһҶОјҡ°ыңyРтЈЁFFPE»тИ«СӘЈ©
- І®әАЙъОп2024ПөБРЕаУ–(xЁҙn)°аЈЁкғОчХҫЈ©ҲуГый_Ҷў
- І®әАҶОјҡ°ыFlexДкЦРҫЮ»ЭЈ¬1.1W/ҳУұҫ»о„УғrјҙҢўҪШЦ№
- Л{ҫ°ҝЖРЕDAP-seqјјРg(shЁҙ)ПакP(guЁЎn)ОДХВ6ЯB°l(fЁЎ)Ј¬ҝӮIF 95.2


Copyright(C) 1998-2025 ЙъОпЖчІДҫW(wЁЈng) лҠФ’Јә021-64166852;13621656896 E-mailЈәinfo@bio-equip.com